Cepheusidae
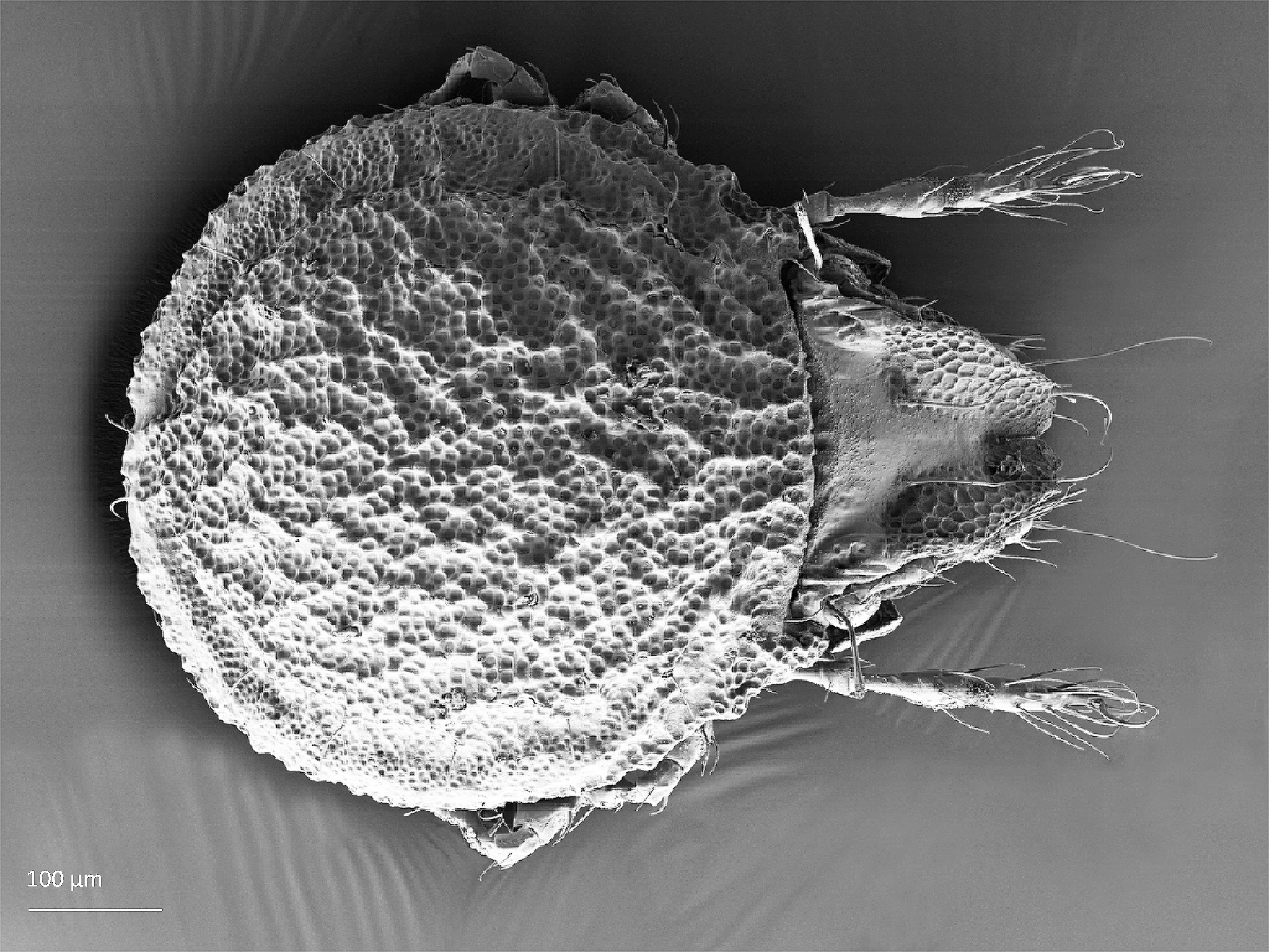
The family Cepheusidae occurs worldwide except in Antarctica. Juveniles bear scalps from previous instars and have peculiar leaf-like setae. They are reputed to be among the most beautiful oribatids known.
- Innhold
- Characteristics
- Habitat and ecology
The family Cepheusidae includes 15 genera, 89 species and 2 subspecies (Subías 2023).
Characteristics
The adults are usually large, seldom medium-sized (500–1200 µm long) and brown or dark brown. The notogaster is relatively broad and has often rounded protrusions in the front. The notogaster has 9–10 pairs of setae, all these setae are located in a marginal or posteromarginal position, while the central part of the notogaster lacks setae. The body surface is often sculptured, and it is smooth only in Conoppia species. Juveniles bear scalps from previous instars and have peculiar leaf-like setae (Weigmann 2006, Norton and Behan-Pelletier 2009, Behan-Pelletier and Lindo 2023).
Habitat and ecology
Cepheusidae species are found in forests, where they are common in moss and litter. They can also be found under lichens, on fruiting bodies of fungi on trees, and in rotting wood (Norton and Behan-Pelletier 2009, Behan-Pelletier and Lindo 2023). Cepheus latus (C.L. Koch, 1835) is a true wood borer in birch twigs (Wallwork 1958) and is macrophytophagous (Mitchell 1978), which means that it feeds on higher plant material only. Fossil Cepheus species are known from Baltic amber (Labandeira et al. 1997).
Cepheus cepheiformis (Nicolet, 1855) described here, is an intermediate host of tapeworms (Denegri 1993). Cepheusidae reproduce sexually (Maraun et al. 2019). The time of the development is known only for Conoppia palmicincta (Michael, 1880). It lasted 375 days from egg to adult under laboratory conditions at 25° C (Pfingstl and Schatz 2021).
References
Behan-Pelletier VM and Lindo Z (2023). Oribatid Mites. Biodiversity, Taxonomy and Ecology. CRC Press, 508 pp.
Denegri GM (1993). Review of oribatid mites as intermediate hosts of tapeworms of the Anoplocephalidae. Experimental and Applied Acarology 17, 567–580. doi.org/10.1007/BF00053486
Labandeira CC, Phillips TL and Norton RA (1997). Oribatid mites and the decomposition of plant tissues in Paleozoic coal-swamp forests. Palaios 12(4), 319–353.
Maraun M, Caruso T, Hense J, Lehmitz L, Mumladze L, Murvanidze M, Nae I, Schulz J, Seniczak A and Scheu S (2019). Parthenogenetic vs. sexual reproduction in oribatid mite communities. Ecology and Evolution 9(12), 7324–7332. doi.org/10.1002/ece3.5303
Mitchell MJ (1978). Vertical and horizontal distribution of oribatid mites (Acari: Cryptostigmata) in an aspen woodland soil. Ecology 593, 516–525.
Norton RA and Behan-Pelletier VM (2009). Suborder Oribatida. In: GW Krantz and DE Walter (eds.). A manual of Acarology, 3rd ed. Texas Tech. University Press Lubbock, 430–564.
Pfingstl T and Schatz H (2021). A survey of lifespans in Oribatida excluding Astigmata (Acari). Zoosymposia 20, 007–027. doi.org/10.11646/zoosymposia.20.1.4
Subías LS (2023). Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758−2002). Graellsia 2004, 60 (número extraordinario), 3−305. Updated 2023 – 18 actualization, 540 pp., access December, 2023.
Wallwork JA (1958). Notes on the feeding behaviour of some forest soil Acarina. Oikos 9, 260–271.
Weigmann G (2006). Hornmilben (Oribatida). Die Tierwelt Deutschlands. 520 pp. Vol. 76, Goecke and Evers, Keltern.