Torvmoseslekta
Sphagnum





Vår torvmoseflora har et enestående artsmangfold og rommer alle de europeiske torvmosene med unntak av fire arter som også er inkludert her. Torvmosene i Norge teller så langt i alt 57 arter med underarter. Noen arter som ikke er funnet hverken i Norge eller Europa så langt er også med for å stimulere til nyfunn. Alt i alt er derfor 65 torvmoser avbildet og nærmere omtalt.
- Innhold
- Morfologi
- Livssyklus
- Klassifisering, taksonomi og fylogeni
- Klassifisering av torvmoser
- De norske torvmosene, arter og underarter
- Utbredelsesgrupper
- Siden siteres som:
Det mest iøynefallende trekket som skiller torvmoser fra andre moser er at skuddene mangler rotaktige festeorganer. Andre særegne trekk er at stengelen har tettstilte greinknipper og bladene består av to typer celler, store døde vannceller og mindre og smalere fotosyntetiserende klorofyllceller. De runde sporehusene sitter på en falsk stilk (pseudopodium) i toppen av moseskuddet, og åpner seg og sprer sporene eksplosjonsaktig om høsten.

Trøndertorvmose er vår mest særegne art, bare funnet i Trøndelag i hele verden. Men spennende er også litorvmose S. rubiginosum og sylfidetorvmose S. venustum som i Europa også bare er kjent fra Norge så langt. Fire av torvmosene er bare funnet på Svalbard i Norge, og er et fascinerende arktisk innslag i vår torvmoseflora.
Morfologi
Gametoforen

Gametoforoppbygning hos sumptorvmose med stengel, greinknipper og hode. Illustrasjon fra Schimper (1875) (t.v.); gametoforskuddet har også to sporofytter på pseudopodier. Foto av hannlig gametofor (t.h.).
Gametoforskuddet består av en ± opprett stengel som mangler rotaktige festeorganer (rhizoider) som vi finner hos andre moser. I toppen av skuddet sitter et apikalt vekstpunkt som sørger for ± kontinuerlig lengdevekst i vekstsesongen, mens skuddet suksessivt dør i den andre enden. På stengelen finner en mer eller mindre tettstilte greinknipper (fascicler). Hvert greinknippe består oftest av 2–3 utstående greiner og 1–3 spedere hengegreiner orientert nedover langs stengelen. De utstående greinene har oftest 30–50 tettstilte og taklagte greinblad. Videre finner en langs stengelen spredtstilt stengelblad. Toppen av moseskuddet med vekstpunktet og tett sammenstilte unge greiner og bladanlegg kalles for hodet (capitulum). Den aller innerste delen av hodet med vekstpunktet og unge grein- og bladanlegg er utformet som en knoppaktig dannelse (toppknoppen) som er synlig i varierende grad.
Bladbygning
Greinbladene sitter tett sammenstilt på greinene, mens stengelbladene sitter spredd på stengelen i en 2/5-spiral. Grein- og stengelbladene har oftest ulik form. Bladene er ét cellelag tykke og mangler bladnerve. Bladene er bygd opp av et nettverk av to typer celler:
- smale, levende klorofyllceller som står for fotosyntesen
- store, pølseformede, tomme hyalinceller (vannceller) som har en sentral rolle i vannopptak og vannlagring
Hyalincellene hos greinbladene har normalt spiralaktige forsterkningslister (fibriller) på sine innervegger, mens slike mangler på stengelblad hos mange arter. Fibrillene bidrar til at cellene bedre kan opprettholde sin form under tørke. I hyalincellenes vegger – særlig på greinbladene – finner en som regel hull (porer) i varierende antall og av varierende størrelse og utforming. Disse har en funksjon i vannopptaket. For mikroskopiske studier er det nødvendig å farge bladpreparatene for å få fram porestrukturen på en oversiktlig måte.
Sporofytten
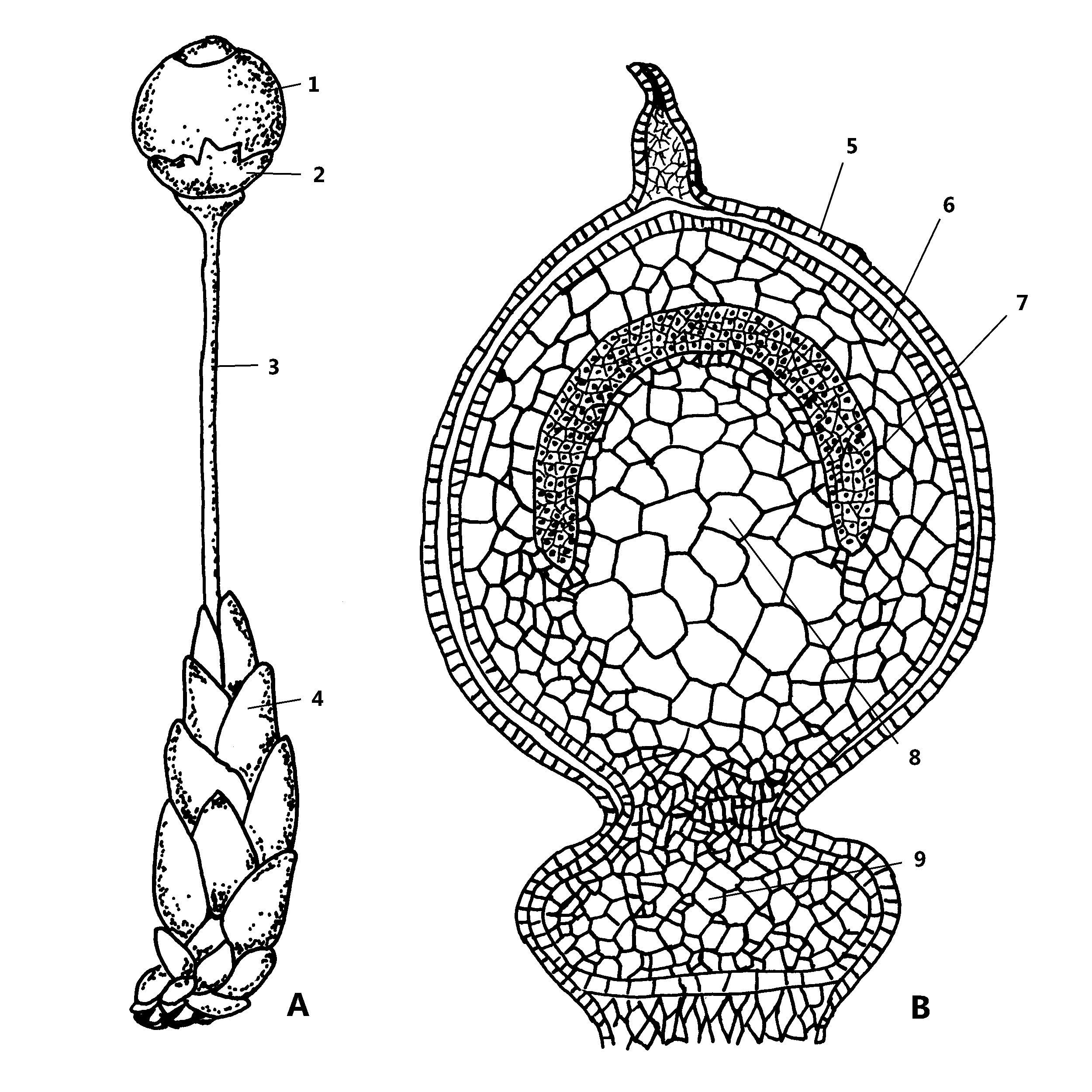
Sporofytt hos torvmoser. Modent sporehus på pseudopodium (A). Skjematisk lengdesnitt gjennom ung sporofytt (B). 1: Sporehus med lokk. 2: Rest etter arkegonieveggen (vaginula). 3: Pseudopodium. 4: Perichaetialblad. 5: Arkegonieveggen. 6: Sporehusveggen. 7: Arkesporiet. 8: Kolumella. 9: Sporehusfoten.
Sporofytten mangler sporehusstilk (seta) til forskjell fra andre bladmoser, og det er bare selve sporehuset som er synlig utenfra. Sporehuset er rundaktig, grønt og fotosyntetiserende i ungt stadium, oftest varierende brunt i moden tilstand. Det sitter festet til gametoforen gjennom en fot, og får det meste av sin næring fra gametoforen gjennom opptak fra denne. Sporehuset har på toppen et lokk (operculum) som åpner seg eksplosjonsaktig ved sporespredningen. Før sporehuset er modent, dekker restene av arkegonieveggen sporehuset som en tynn hinne (vaginula). Når sporehuset er modent, strekker festepunktet til gametoforen stilkaktig på seg til et pseudopodium. Dette har samme funksjon som sporehusstilken hos andre bladmoser, men er haploid og en del av gametoforen, ikke sporofytten. På pseudopodiet sitter det egne blad – såkalte perichaetialblad – som avviker fra både grein- og stengelbladene i morfologi. Sporehuset er omgitt av en kapselvegg som inneslutter det sporedannende vevet (arkesporiet) og en midtstilt, halvkuleformet søyle (kolumella). En fot forankrer sporehuset i pseudopodiet.
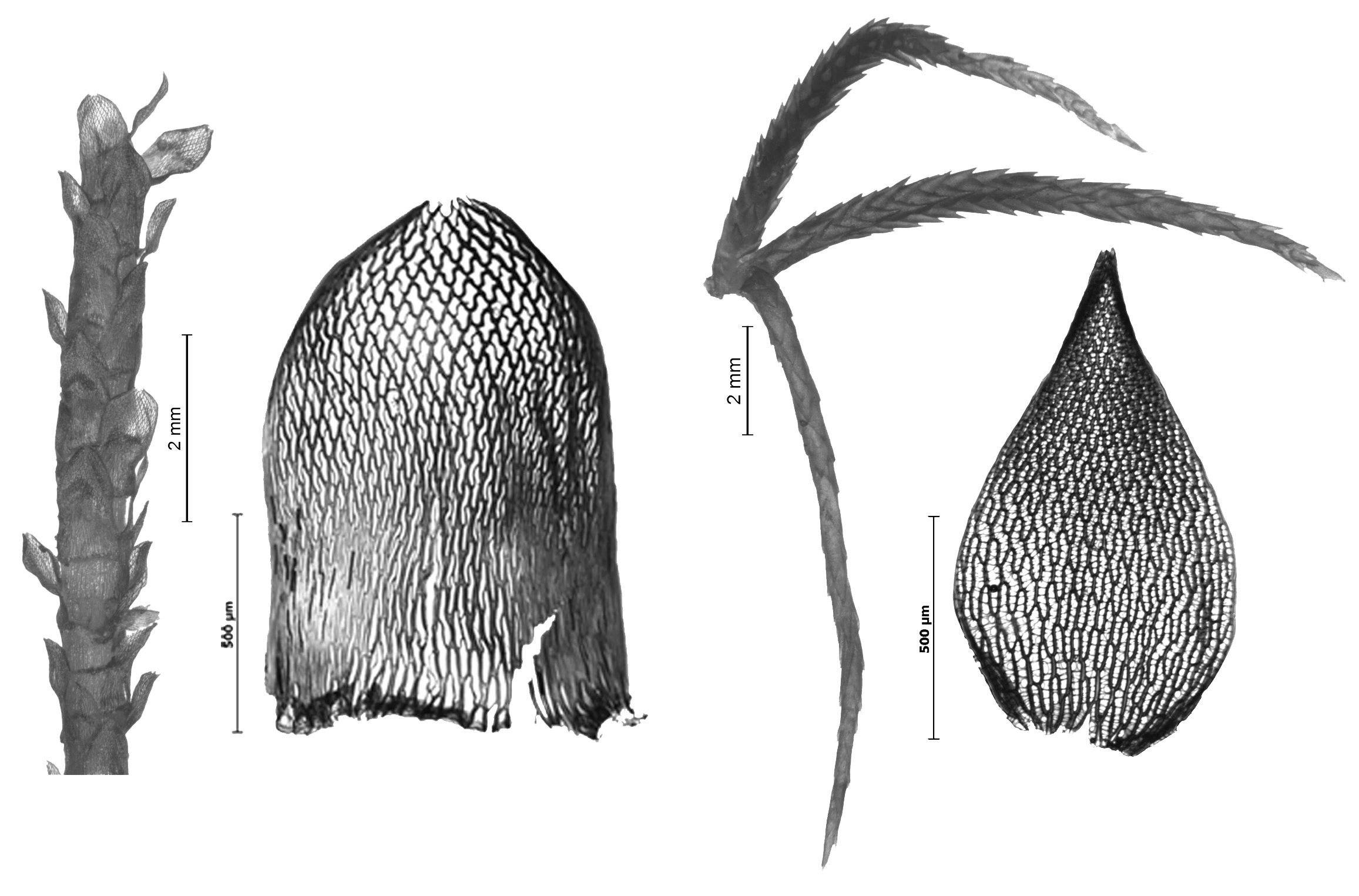
Tvaretorvmose Sphagnum russowii. Fra venstre: Utsnitt av stengel med oppoverrettede stengelblad. Stengelblad. Greinknippe med to utstående greiner og en hengegrein. Greinblad fra midten av ustående grein.

Livssyklus
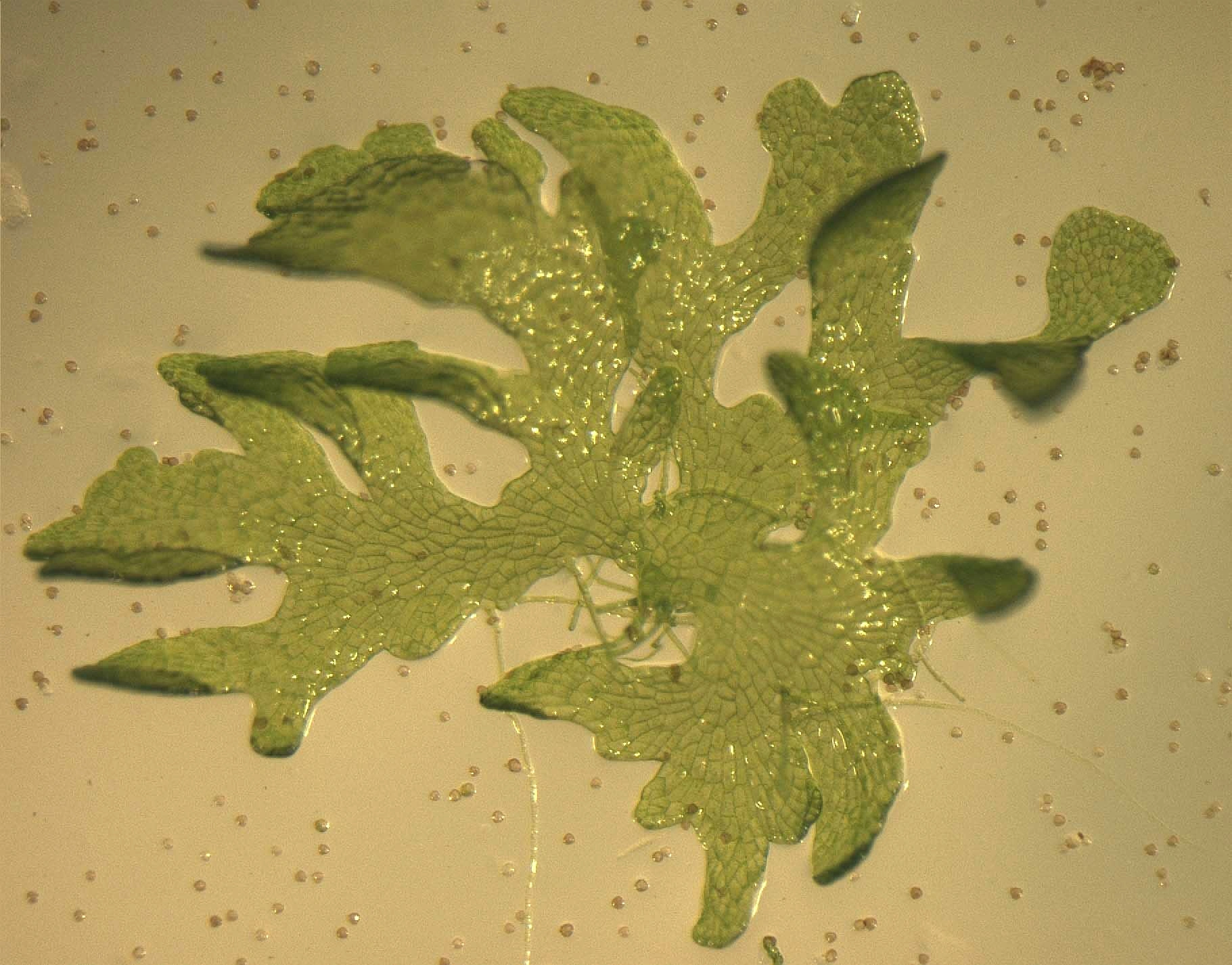
Bladformet protonema og små sporer som ikke har spiret.
Torvmosene har generasjonsveksling mellom en haploid generasjon (gametofyttgenerasjonen) og en diploid generasjon (sporofyttgenerasjonen). Gametofyttgenerasjonen består i første fase av et grønt, først trådaktig, til slutt bladaktig (thalloid) protonema (flertall: protonemata) forsynt med rotaktige festeorganer (rhizoider). Protonemata ligger i markoverflaten og spirer normalt opp fra sporer. De er bare noen få millimeter store og vanskelige å oppdage i felt. På protonemaet dannes et eneste skudd, en gametofor. Gametoforen utgjør den fotosyntetiserende moseplanten vi ser ute, og mange gametoforer opptrer som regel tett sammenstilte i matter og puter av varierende størrelse. Protonemaet og gametoforen utgjør gametofytten. Gametoforen danner hannlige og hunnlige kjønnsorganer (antheridier og arkegonier). De hannlige kjønnscellene (spermatozoidene) er selvbevegelige ved hjelp av to flageller, og overføringen fra antheridiene til arkegoniene forutsetter vann. Det befruktede egget i arkegoniet (zygoten) utvikler en diploid sporofytt som blir sittende festet til gametoforen gjennom en fot (pseudopodium). Etter reduksjonsdelingen (meiosen) i sporehuset dannes tallrike, haploide sporer. Sporene sprees og gir ved spiring på egnet voksesubstrat nye protonemata med gametoforer. Gametoforen (moseplanten) er flerårig, mens sporofytten er ettårig og dør etter sporeslippet.

Fra venstre: A: Hunnskudd hos abelstorvmose S. divinum med tre rød-aktige perichaetier blant unge greiner som omgir toppknoppen. B: Arkegonium klar for befruktning. C: Befruktet arkegonium (til venstre) som har utviklet ung sporofytt i det oppsvulmede nedre partiet. D: Unge sporofytter omgitt av rester av arkegoniet og perichaetialblad. E: Gametoforskudd med sporofytter på pseudopodier. Modent, uåpnet sporehus (t. v.), to sporehus som har åpnet seg og sluppet sporene (t. h.).
Klassifisering, taksonomi og fylogeni

Flatbergium novo-caledoniae i familien Flatbergiaceae er kun kjent fra Ny-Caledonia i Stillehavet.
Et omfattende genetisk arbeid med bruk av ulike DNA-baserte metoder har blitt utført av J.A. Shaw og hans medarbeidere ved Duke University i North Carolina fra begynnelsen av 2000-tallet og utover. Dette har ført til grunnleggende ny kunnskap om torvmosenes fylogeni og slektskapsforhold. Den overordnede klassifiseringen og taksonomien av torvmoser i som er anvendt her, følger Shaw et al. (2015), og som avviker en del fra Norges torvmoser (Flatberg 2013), som i hovedsak bygger på Shaw et al. (2010b).
Sphagnopsida (torvmoser i vid forstand) er en gammel organismegruppe på jorda. Den ble trolig spaltet ut fra andre bladmoser for et sted mellom 129 og 319 millioner år siden (Shaw et al. 2010a, b), som indikert gjennom molekylærgenetiske studier. Torvmoser sammen med bladmoseslekten Takakia (med to sjeldne nålevende arter funnet i vestlige Nord-Amerika og Sentral- og Sør-Asia) kan være «søster» til alle andre bladmoser (Chang & Graham 2011). Det eldste fossile funn av torvmoselignende blad hos den utdødde slekten Protosphagnum går tilbake til Perm-tiden (ca. 300–250 mill. år siden). Torvmosenes opprinnelse er trolig knyttet til den sørlige halvkule og den fordelingen av kontinenter og landmasser en hadde da. De tre familiene Ambuchananiaceae, Eosphagnaceae og Flatbergiaceae kan være relikter fra den opprinnelige torvmosefloraen. Denne floraen kan ha vært forholdsvis artsrik, men mangel på fossilt materiale gjør dette til en gjetning. Familien Sphagnaceae skilte lag fra denne gruppen for et sted mellom 34 og 104 millioner år siden. Det store artsmangfoldet av Sphagnum-arter slik vi kjenner det i dag på den nordlige halvkule, har trolig sine røtter i siste del av miocen tidsperiode i tertiærtiden for mellom 7 og 20 millioner år siden (Shaw et al. 2010a). Det økte artsmangfoldet var et resultat av et kaldere klima med dannelse av boreal vegetasjon og myr. Også mange av de torvmoseartene som i dag finnes på den sørlige halvkule, kan ha sin opprinnelse i denne boreale «poolen» av arter på den nordlige halvkule.
Familiene Ambuchananiaceae, Eosphagnaceae og Flatbergiaceae – som bare er kjent fra den sørlige halvkule – inneholder bare fire kjente arter, familien Sphagnaceae de øvrige torvmoseartene. Det er bare Sphagnaceae med den ene slekten Sphagnum som er representanter på den nordlige halvkule i dag. Totalt antall arter i slekten i verdenssammenheng er ikke kjent, da det knytter seg stor usikkerhet til den taksonomiske identiteten til mange av de tallrike artene som tidligere er blitt beskrevet. Særlig gjelder dette de tropiske artene som er beskrevet av den tyske torvmoseforskeren C. Warnstorf mot slutten av 1800-tallet og begynnelsen av 1900-tallet, og som inngikk i hans verdensmonografi «Sphagnologia Universalis» (Warnstorf 1911). Mange av hans arter har vist seg å være voksestedsmodifikasjoner av andre arter, mens andre arter igjen ennå ikke er kritisk vurdert og taksonomisk revidert. Michaelis’ (2011) nye verdensmonografi over slekten omtaler nærmere 300 arter. Dette antallet kan også være noe høyt.
Tidligere klassifiseringer av Sphagnum ble basert på morfologiske bygningstrekk og karakterer. Det har vært tradisjon å dele inn artene som forekommer på den nordlige halvkule, i åtte likestilte hovedgrupper kalt seksjoner. Disse seksjonene tilsvarer i innhold de fem underslektene som er skilt ut her. Seksjonene Polyclada og Insulosa er fremdeles en del av underslekten Acutifolia i den nye klassifiseringen til Shaw et al. (2016), men det nye er at underslekt Squarrosa ikke kan opprettholdes hverken som egen underslekt eller seksjon, men nestes innenfor seksjon Acutifolia. I den nye klassifiseringen er Rigida den basale underslekten og søster til resten av torvmoseslekten. Den har to hovedklader, den ene er søster til underslektene Sphagnum og Acutifolia, den andre til underslektene Subsecunda og Cuspidata.

De norske torvmosene, arter og underarter
Rekkefølgen av arter innenfor de fem underslektene i denne beskrivelsen forsøker å vise kjente systematiske og fylogenetiske relasjoner mellom dem. Men det er foreløpig begrenset med molekylærgenetiske data tilgjengelig for å understøtte de innbyrdes artsrelasjonene, blant annet innenfor den artsrike underslekten Cuspidata. Shaw et al. (2005) gir det fylogenetiske grunnlaget for rekkefølgen av arter innenfor underslekten Acutifolia. Allodiploide hybridarter (se senere) er kjent innenfor underslektene Sphagnum, Subsecunda, Cuspidata og Acutifolia, og mellom underslektene Cuspidata og Subsecunda. Disse kan være vanskelige å plassere i en fylogenetisk sammenheng, ettersom også foreldreforholdene til dels er usikre eller ukjente.
Utbredelsesgrupper
Mange av de norske torvmosene har utbredelser som er klart relatert til utbredelsen av trinn og mønstre innenfor de ulike sonene og seksjonene. Dette gir grunnlag for å klassifisere arter sammen i plantegeografiske grupper (utbredelsesgrupper) der en kan se for seg forholdsvis like responser på makroklimatiske variabler hos arter innenfor hver av gruppene, og ulike responser mellom gruppene. Men andre faktorer kan influere på det utbredelsesbildet en bestemt art viser, for eksempel knyttet til klimahistorie og innvandring etter siste istid, spredningsevne over tid, etablering og konkurranse med andre arter, habitattilgang og menneskelig påvirkning. Med unntak av ubikvistene er utbredelsesgruppene gitt navn etter artenes geografiske tyngdepunkt i forhold til sør-nord, vest-øst og kombinasjoner.
Forklaring på bioklimatiske soner og seksjoner, som nevnes hyppig i dette kapittelet, finner du i lenka under
Ubikvister
Grantorvmose er en vid ubikvist (t.v.), tvaretorvmose er en fastlandsubikvist (t.h.)
Ubikvister omfatter arter som er vidt utbredd gjennom det meste av fastlandet i boreonemoral sone og alle boreale soner og de to sørarktiske tundrasonene i Finnmark, og i alle seksjoner. Alle artene har i tillegg i det minste strøforekomster i lavalpin sone og noen få arter når også mellomalpin. Mange av artene er i tillegg også funnet på Svalbard.
Vide ubikvister
Artene her er vidt utbredd på fastlandet og er i tillegg funnet på Svalbard. Beitetorvmose og spriketorvmose er også funnet på Jan Mayen.
Fastlandsubikvister
Artene her har en vid utbredelse på fastlandet, og er alle med unntak av kroktorvmose Sphagnum subsecundum funnet i de sørarktiske områdene i Finnmark. Ingen av artene er funnet på Svalbard.
Sørlige arter
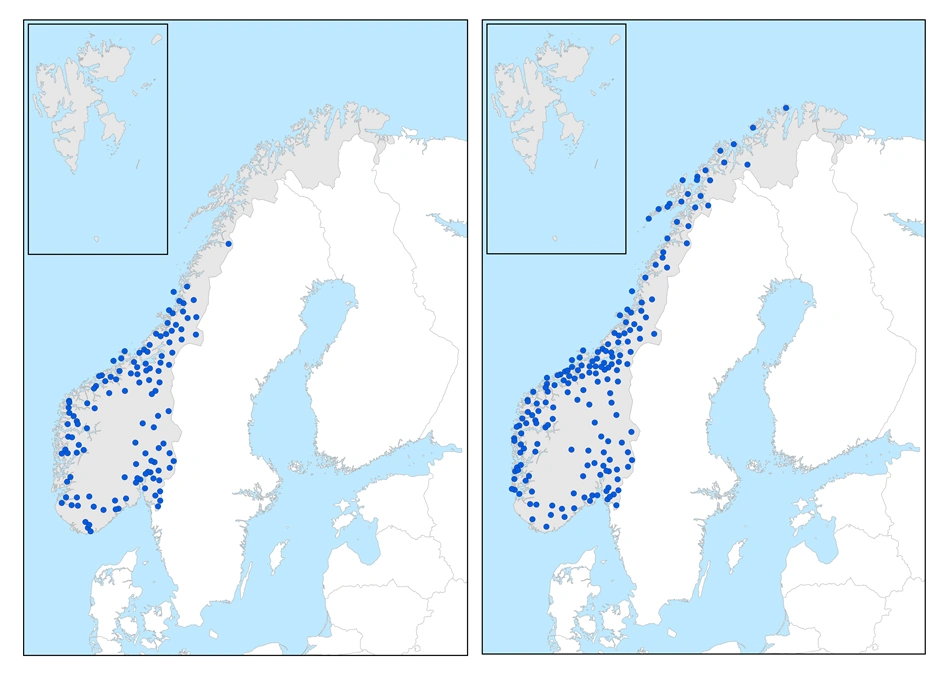
Fagertorvmose (t.v.) er en moderat sørlig art. Rødtorvmose (t.h.) er en svakt sørlig art.
Den sørlige utbredelsesgruppen omfatter arter som er utbredt i alle fastlandssoner til og med nordboreal sone, men som bare mer unntaksvis forekommer over skoggrensen i lavalpin sone, eller nord for skoggrensen inn i sørarktiske områder i Finnmark. De fleste artene har sine nordligste kjente forekomster i nordboreal sone i Finnmark, men bleiktorvmose Sphagnum flexuosum i Troms og fagertorvmose S. pulchrum i Nordland. Artene forekommer i alle seksjoner, men noen kan være sjeldne eller fraværende i svakt kontinental seksjon i Troms og Finnmark, til dels også i Hedmark.
Moderat sørlige arter
I Sør-Norge er fagertorvmose funnet i alle boreale soner og seksjoner, også svakt kontinental seksjon (Hedmark: Femundsmarka). Den har kjent nordgrense i mellomboreal sone/svakt oseanisk seksjon i Nordland. Den har nordgrense på tilsvarende breddegrad i Lule Lappmark på svensk side. Det er vanskelig å finne klimatiske årsaker til at arten ikke er funnet lenger nord.
Svakt sørlige arter
Artene her har sine kjente nordgrenser i nordboreal sone i Nordland, Troms eller Finnmark, og har til dels spredde forekomster i lavalpin sone. De forekommer vanlig til mer spredd i alle øvrige soner under skoggrensen fra boreonemoral til nordboreal sone, og i alle seksjoner.
Sørvestlige arter
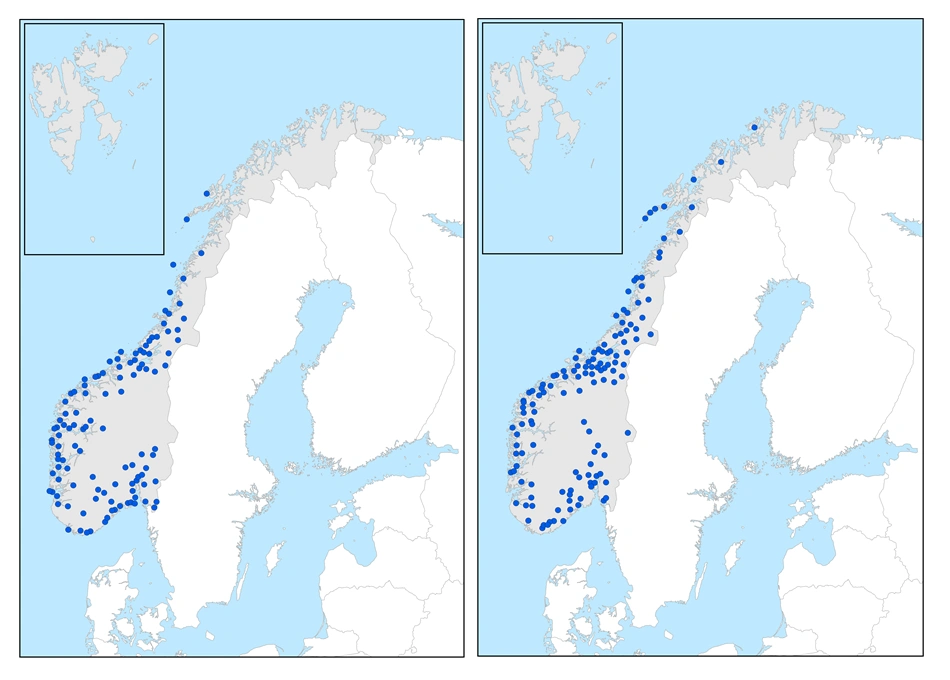
Sumptorvmose (t.v.) er en moderat sørvestlig art. Lyngtorvmose (t.h.) er en svakt sørvestlig art.
De sørvestlige artene har sitt tyngdepunkt i utbredelsen i sterkt oseanisk seksjon → svakt oseanisk seksjon. De kan forekomme i overgangsseksjon og unntaksvis i svakt kontinental seksjon i Sør-Norge. De forekommer alle i boreonemoral og sørboreal sone, mens forekomstsettet i mellomboreal og nordboreal sone varierer. Den kjente nordgrensen langs vestkysten varierer fra Nordland til Finnmark. Nordgrensen på Østlandet og østgrensen i Midt-Norge virker å være korrelert med humiditet/oseanitet, og nordgrensen langs vestkysten med sommertemperatur.
Moderat sørvestlige arter
Disse artene har en forholdsvis vid kystutbredelse fra sterkt oseanisk → svakt oseanisk med utløpere i overgangsseksjon, og har sin kjente nordgrense i mellomboreal sone i sterkt oseanisk seksjon, eller klart oseanisk seksjon i Nordland. Øst- og høydegrensen i Midt-Norge ligger i mellomboreal sone/klart oseanisk seksjon. På Østlandet har en nordgrensen i sørboreal sone(→mellomboreal sone) i svakt oseanisk og overgangsseksjon, men artene mangler i sørboreal sone og mellomboreal sone i svakt kontinental seksjon, de fleste steder også i overgangsseksjon og svakt oseanisk seksjon.
Svakt sørvestlige arter
Artene her har en vid kystutbredelse fra sterkt oseanisk til svakt oseanisk seksjon. De har sin kjente nordgrense i nordboreal sone/svakt oseanisk seksjon i Troms eller Finnmark. I Sør-Norge har artene høydegrense i nordboreal sone, noen steder også med utløpere inn i lavalpin sone. På Østlandet går de nord til mellomboreal sone/svakt oseanisk seksjon, med utløpere inn i svakt kontinental seksjon.
Gruppen er heterogen, og særlig glasstorvmose Sphagnum angermanicum har en avvikende, markert oppsplittet utbredelse.
Vestlige arter
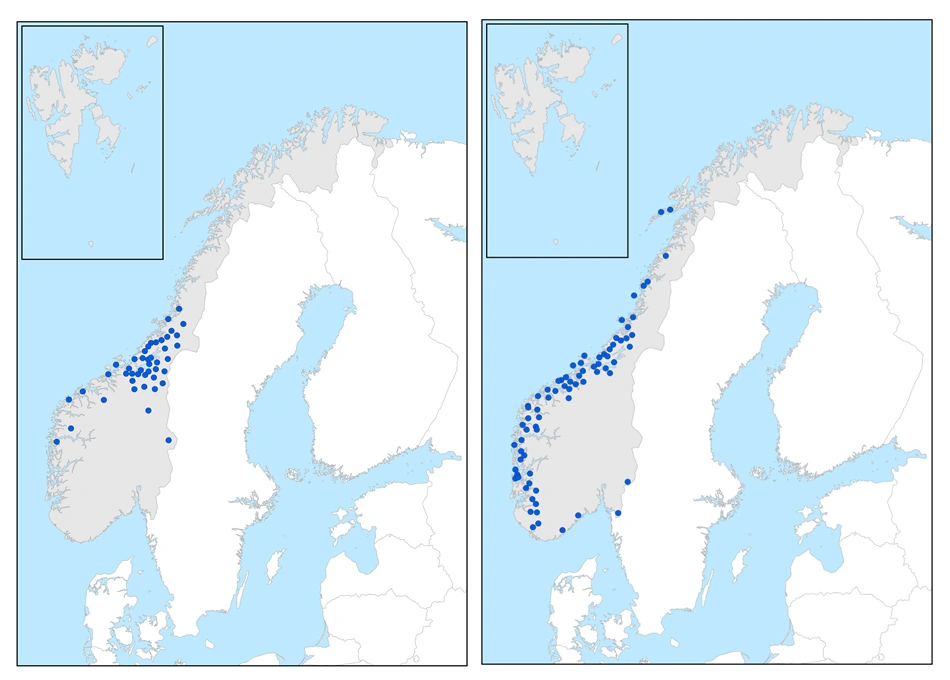
Litorvmose (t.v.) er en midtnorsk, vestlig art. Kysttorvmose (t.h.) er en sterkt til moderat vestlig art.
De vestlige artene er en sammensatt gruppe som har sin utbredelse eller hovedutbredelse knyttet til sterkt oseanisk og klart oseanisk seksjon, og som med få unntak er helt fraværende fra overgangsseksjon og svakt kontinental seksjon. Den sonale tilhørigheten varierer.
Midtnorske, vestlige arter
Artene her har hovedutbredelse i sørboreal og/eller mellomboreal sone i klart oseanisk seksjon i Midt-Norge, med individuelle særtrekk i utbredelsen. De er i Europa ikke funnet utenom Norge, og trøndertorvmose og sylfidetorvmose er bare funnet nord i Trøndelag. Utbredelsesbildet er vanskelig å forklare ut fra dagens klimaforhold og kan være et resultat av historiske, innvandringsmessige forhold. Særlig sylfidetorvmose kan også være oversett, og nyfunn kan endre utbredelsesbildet.
Sterkt til moderat vestlige arter, en heterogen gruppe
Artene har sin utbredelse eller hovedutbredelse i sterkt oseanisk seksjon, men forekommer også i klart oseanisk seksjon og unntaksvis i svakt oseanisk seksjon. Artene mangler i det store og hele på Østlandet, med unntak av særlig Østfold. Gruppen er heterogen, og artene viser individuelle mønstre, særlig når det gjelder sonal utbredelse. Utbredelsesbildet kan derfor være relatert til ulike variabler knyttet opp mot kystklima. Horntorvmose og bruntorvmose har usikker utbredelse.
Østlige arter
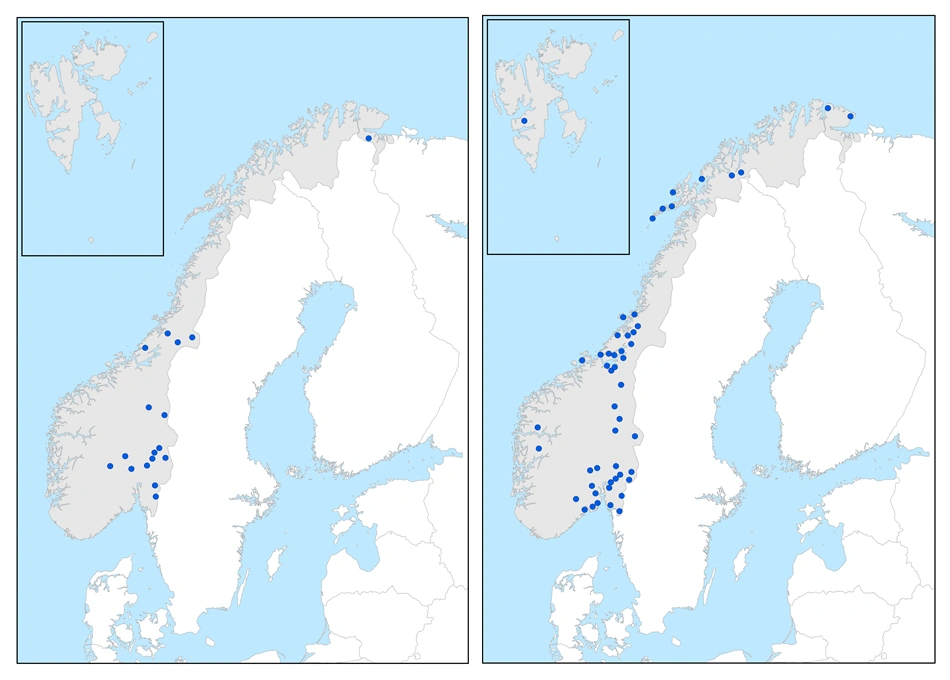
Huldretorvmose (t.v.) er en moderat østlig art. Butt-torvmose (t.h.) er en svakt østlig art.
Østlige arter har hovedtyngden av forekomstene i svakt kontinental seksjon og overgangsseksjon, men har vestlige utkantforekomster både i sterkt oseanisk og klart oseanisk seksjon. De har ingen klar sonetilknytning, men er ikke funnet over skoggrensen. Det totale spennet i utbredelse med spredde utkantlokaliteter i sterkt oseanisk seksjon langs vestkysten kan indikere at dagens utbredelse til dels har historiske årsaker, og at artene her har hatt en forholdsvis sen innvandring etter siste istid fra øst. Alle artene ført hit er funnet med sporehus i Norge, men forholdsvis sjelden. Bare gråtorvmose S. majus subsp. norvegicum er funnet på de britiske øyer.
Moderat østlige arter
Denne arten har et klart østlig tyngdepunkt i svakt kontinental seksjon og overgangsseksjon, men er også funnet med en vestlig utkantforekomst i sterkt oseanisk seksjon på vestkysten sør i Trøndelag: Åfjord. Den har ingen kjente norske forekomster mellom Trøndelag og Finnmark: Sør-Varanger, men forekommer på tilsvarende strekning på svensk side.
Svakt østlige arter
De to første har spredde vestkystforekomster i sørboreal og mellomboreal sone i sterkt oseansik seksjon fra Trøndelag og nordover, men synes å mangle eller er sjeldne i sørboreal og boreonemoral kystsone lenger sør på vest- og sørvestkysten. Butt-torvmose er også funnet i mellomarktisk tundrasone på Svalbard.
Nordlige og nordøstlige arter
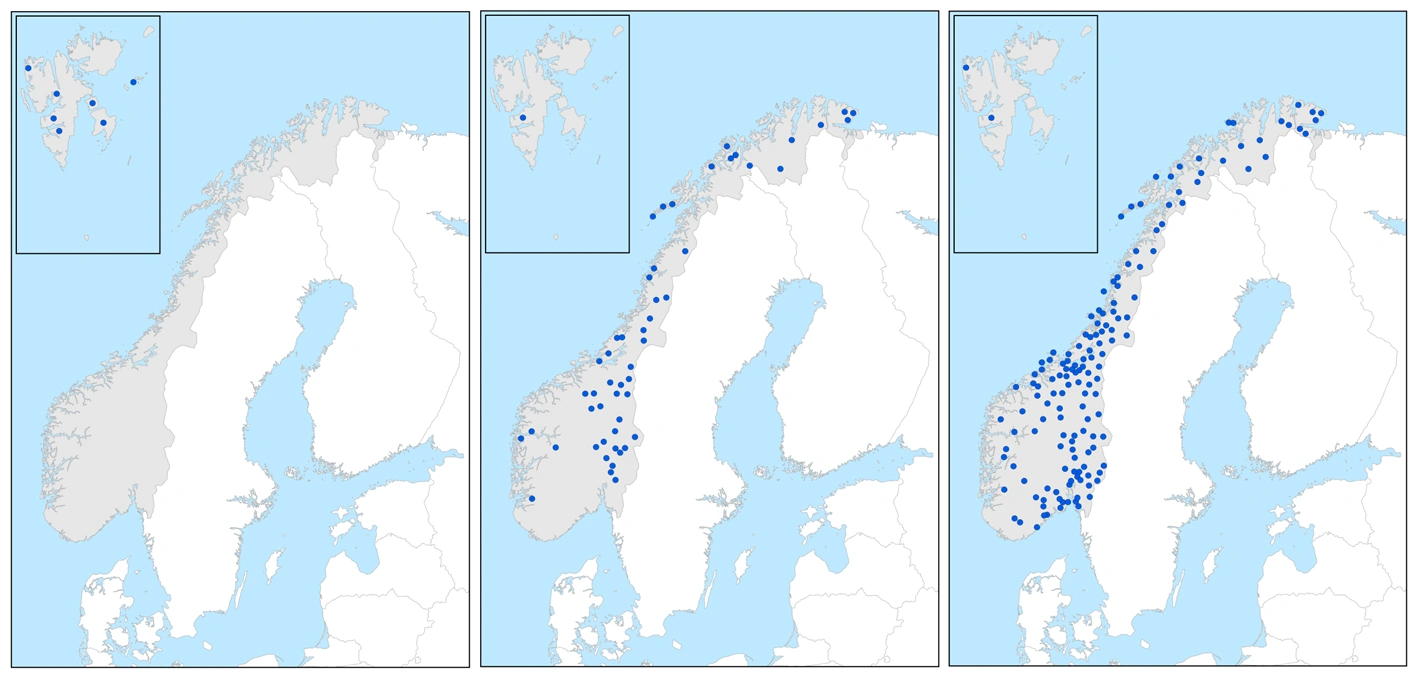
Istorvmose(t.v.) er en sterkt nordlig art, fjelltorvmose (midten) er en moderat nordlig art og svelttorvmose (t.h.) er en svakt nordlig/nordøstlig art.
Artene i den nordlige og nordøstlige utbredelsesgruppen er bare funnet på Svalbard, på Svalbard og fastlandet med en nordlig trend i utbredelsen, eller bare på fastlandet med et nordøstlig tyngdepunkt.
Sterkt nordlige arter
Disse artene er bare funnet på Svalbard i mellomarktisk tundrasone→(nordarktisk tundrasone)/ svakt kontinental seksjon→(overgangsseksjon) og ikke på fastlandet. I arktiske områder utenom Svalbard (Grønland, Kanada, Alaska, Russland) er disse artene til dels også funnet i arktisk kratt-tundrasone og sørarktisk tundrasone. En kan derfor ikke utelukke forekomst av en eller flere av disse i sørarktiske deler av Finnmark. Men de kan også mangle der av historiske og ikke klimatiske årsaker.
Moderat nordlige arter
Denne arten er funnet på Svalbard og fastlandet. På fastlandet har den spredd utbredelse med hovedtyngde i mellomboreal sone→nordboreal sone/overgangsseksjon→svakt kontinental seksjon, og inngår i sørarktisk tundrasone/overgangsseksjon i Finnmark. Det er utkantforekomster i mer oseaniske seksjoner. Arten er ikke funnet på De britiske øyer.
Svakt nordlige/nordøstlige arter
Dette er arter som på fastlandet er vanlige/hyppige i mellomboreal og nordboreal sone, mindre frekvente i sørboreal og boreonemoral sone, eller sjeldne eller ikke funnet i boreonemoral sone. De opptrer spredd til sjelden i sterkt oseanisk seksjon og har hovedutbredelsen i svakt oseanisk seksjon, overgangsseksjon og svakt kontisnental seksjon. Bjørnetorvmose Sphagnum lindbergii og svelttorvmose S. balticum er funnet på Svalbard, mens de øvrige bare er funnet på fastlandet. Alle artene med unntak av lapptorvmose er funnet i sørarktiske områder i Finnmark. Dette er arter/underart som mangler eller er sjeldne på De britiske øyer.

Siden siteres som:
Flatberg KI (2015). Torvmoseslekta Sphagnum L. www.artsdatabanken.no/Pages/192053. Nedlastet <dag/måned/år>