Muslingkreps
Ostracoda

Hvitmuslingkreps Candona candida.
Muslingkreps (Crustacea, klasse Ostracoda) er en gruppe små krepsdyr som lever både i havet og i ferskvann. Det norske navnet henspiller på den pussige morfologien, som er så lik en musling at man ved første øyekast kunne forveksle dem.
- Innhold
- Bakgrunn
- Taksonomi
- Å finne muslingkreps
- Preparering - noen praktiske råd
- Observasjoner under lupen
- Preparering og mikroskopiske kjennetegn
Bakgrunn
Som mollusker har muslingkreps et todelt, kalsifisert skall, som kan lukkes ved hjelp av et knippe tverrgående muskler og et hengselsystem på ryggsiden. Også skallenes omriss ligner ofte på muslinger og skjell. Anatomien avslører dem imidlertid straks som ekte krepsdyr. Bak skallet skjuler det seg både bein, munndeler og flere andre organer som lett kan relateres til hummer, krabber og kreps. Historisk er muslingkreps svært gamle, og fordi de produserer mineraliserte skall som lett blir bevart som fossiler og ofte opptrer i store mengder, er de, i likhet med foraminiferer og kiselalger, viktige ledefossiler.
De opprinnelige muslingkrepsene var bunnlevende saltvannsformer, men allerede tidlig ble mange arter pelagiske, og enkelte tok også veien via brakkvannspytter i tidevannssonen og over i ferskvann. Fortsatt er diversiteten av muslingkreps med 13000 arter mye høyere i havet, men anslagsvis 2000 arter er knyttet til ferskvann. I Europa er muslingkrepsfaunaen i ferskvann helt dominert av én bestemt overfamilie, Cypridoidea, som i sin tur deles inn i fire familier. To av disse - Candonidae og Cyprididae - er særlig artsrike og viktige, og har flere tydelige karakteristika, som i det følgende vil få det meste av oppmerksomheten.
I Norge ble muslingkreps for første gang kartlagt av Georg Ossian Sars (1837-1927), og han beskrev også en rekke nye arter. Særlig grundig undersøkte han området omkring Oslofjorden, men han arbeidet også omkring Bergen, og foretok reiser opp gjennom Gudbrandsdalen og Trøndelag. Resultatet av dette var en første oversikt over 56 norske arter. I Sverige foretok Gunnar Alm (1889-1962) et tilsvarende kartleggingsarbeid (Alm 1915).
Etter Sars ble det en pause i interessen for muslingkreps her i landet. Etter hvert ble andre krepsdyr, særlig cladocerer og copepoder, viktige forskningsfelt for både økologiske og molekylære studier. Mye forskning rettet seg mot livet i de åpne vannmassene, der limnologer allerede hadde utviklet mye relevant kunnskap. Muslingkreps, som er helt eller delvis bunnlevende, gikk ofte «under radaren» - eller rettere sagt under planktonhåvene som trålet norske innsjøer på kryss og tvers gjennom det 20. århundre.
Ikke desto mindre har muslingkreps stor betydning for næringskjeder og produksjon av biomasse i mange innsjøer, tjern og dammer. I virkeligheten finnes det ikke noe skarpt skille mellom planktoniske og bunnlevende samfunn. De aller fleste akvatiske dyr nyttiggjør seg ressurser begge steder. I enkelte systemer kan tetthetene av muslingkreps bli svært høye, og de er utvilsomt viktige i mange ferskvannssystemer, ikke minst som føde for fiskeyngel.

Fiplingdalen våtmark ved Børgefjell, et interessant område med mye små myrkulper, dammer og tjern, og en større elv, har ganske høy diversitet.
Taksonomi
I europeiske ferskvann er det altså registrert tre distinkt ulike taxa (overfamilier) av muslingkreps: Darwinuloidea, Cytheroidea og Cypridoidea. Sistnevnte er uten sammenligning den største gruppen. Førstnevnte har så vidt man vet kun én art i Norge – Darwinula stevensoni. Det er funnet noen flere arter av Cytheroidea (Sars 1928), men kun en art – Limnocythere inopinata - er mer utbredt. Begge de nevnte artene har trekk som gjør at de lett identifiseres.
Det som da står igjen er den artsrike familien Cypridoidea, som i det følgende skal stå i sentrum. Familien inneholder flere underfamilier.
Å finne muslingkreps

Saltpåvirkede rock pools rett ved havet i Salten, der er det ofte særegne samfunn av muslingkreps.
Fordi så få tok seg tid til å sette seg inn i denne organismegruppen siden Sars’ dager fikk muslingkreps etter hvert rykte på seg for å være vanskelige å bestemme. Men som også faktaarkene og nøkkelen (kommer) viser, er det slett ikke riktig. Så sant man har tilgang på en stereolupe lar mange arter seg bestemme umiddelbart ut fra form, størrelse, farger og ornamentering. Om man går videre vil man ha uvurderlig nytte av Claude Meisch (2000) sin storartede monografi med nøkkel og veiledning, og med fyldig gjennomgang av alle europeiske arter (riktignok er ikke Svalbard inkludert, der et par karakteristiske arter, særlig praktmuslingkreps Tonnacypris glacialis og Fabaeformiscandona reticulata, er vanlige). Etter at NIVA i regi av Artsdatabanken foretok en landsdekkende kartlegging er nå 62 arter av muslingkreps registrert i norske ferskvannssystemer.
En annen grunn til at muslingkreps lenge gikk under biologenes radar er at de ofte har høyest mangfold og tettheter i miljøer som planktonbiologer ikke har oppsøkt like hyppig. Man kan for eksempel finne ganske diverse samfunn av muslingkreps i grøfter langs landeveier eller i åkerkanter, i små dammer, vannhull, pytter eller i skogstjern. Småtjern rundt Oslofjorden hadde høyest diversitet, med opp til 9 ulike arter (Tjernsrud-tjern og Snarøytjern i Bærum). En egen fauna finnes i såkalte rock pools, dvs saltpåvirkede regnvannspytter på svabergene langs kysten. Og flomdammene langs de store elvene, som i Tana, Målselv, langs Flisa eller i Gudbrandsdalslågen, kan også by på interessante funn (for eksempel Pseudocandona albicans). Enkelte arter har en forkjærlighet for kilder og oppkommer i fjellheimen (som bekkemuslingkreps Potamocypris pallida), ofte også i slam som bare så vidt er dekket av vann. I myr kan de også leve i fuktig torvmose. Andre arter lever i bunnsedimentene på større dyp i store innsjøer, eller mellom småstein på bunnen av stilleflytende elver. Egenartede samfunn av muslingkreps fant vi i varme kilder på nordsiden av Svalbard (Heterocypris salina, Fabaeformiscandona reticluata), og i kalksteinsgrotter (Cryptocandona vavrai, dvergmuslingkreps Nannocandona faba).
Skal man gjøre seg kjent med denne fascinerende organismegruppen, må man altså ikke bare ta prøver fra innsjøer og tjern, men også oppsøke andre små vannmiljøer av ulikt slag.
Også selve prøvetakingen bør tilrettelegges noe. Det er lite effektivt å bruke vanlig planktonhåv, fordi de fleste muslingkreps lever på eller nede i bunnsubstratet. Skal man finne dem må man målrettet prøveta dette miljøet. Det gjøres ved simpelthen å tråkke varsomt omring i vannet så litt av bunnsedimentet virvles opp, og så øse 20-30 liter vann gjennom et 250 eller 500 µm planktonfilter ved hjelp av et litermål. Om man til slutt klemmer filter med innhold sammen blir man kvitt overflødig vann, og resten kan fikseres, helst på etanol, fordi fargene dermed også bevares. Veigrøfter, pytter og kilder med lite vann kan prøvetas ved at man slår forsiktig i vannfilmen og filtrerer det man får tak i. Om man har en hvit plastboks, for eksempel en isboks, kan man sjekke filtratet før fiksering, og se hvordan de forskjellige artene beveger seg.

I innlandet i Sør-Norge, som her i Flå i Hallingdal, finnes masse små dammer, myrtjern og bekkefar, der muslingkreps trives bra.
Preparering - noen praktiske råd
Faktaarkene gir en presentasjon av de vanligste artene her i landet, samt av en håndfull arter som av andre grunner kvalifiserer til spesiell omtale. Generelt har vi valgt ut arter der fotografier gir hjelp til bestemmelsen. En god del muslingkreps bør slik kunne identifiseres ved form, farge, ornamentering, behåring og størrelse, ved hjelp av faktaarkene. For en del arter, og ikke minst den store familien Candonidae, er imidlertid preparering og mikroskopi nødvendig. Nøkkelen leder vei til de vanligste norske artene, og knytter an til Sars (1928) sin oversikt over norsk ostracode-fauna, og til Langelands nøkkel (Langeland 1973). Men om man vil gjøre seg mer grundig kjent med muslingkreps trenger man en komplett nøkkel. En slik nøkkel finnes som nevnt i Claude Meisch sin svært grundige og solide monografi (Meisch 2000).
Det beste er å arbeide med 5-6 individer samtidig. Første skritt er å avgjøre om prøven inneholder voksne (adulte) individer (juvenile lar seg sjeldent sikkert bestemme). Juvenile er mindre enn 0,5 mm (men se opp for dvergmuslingkreps Nannocandona faba, som også er mindre enn 0,5 mm!), og har gjerne gjennomskinnelige skall, som er svakere mineralisert enn hos utvokste. Med litt øvelse kan man allerede ut fra hastigheten de synker til bunns i petriskålen avgjøre om de er utvokste (adulte har mer kalsium i skallene og synker raskere). Hos utvokste vil man også ofte kunne se det karakteristiske båndet av modnende egg rett under skallet (synlig på bildet av nettmuslingkreps Brandleystrandesia reticulata). Hannenes spermatozoer sees som liknende, men parallelt liggende bånd, side om side i «varv» under skallet. Hanner er imidlertid slett ikke alltid til stede i populasjonen.
Reproduksjonen hos muslingkreps er ofte partenogenetisk, dvs at hunnene har egg som utvikler seg til nye muslingkreps uten befruktning. I mange populasjoner og arter kjenner man bare hunner. I andre populasjoner er imidlertid også hanner vanlige, og muslingkreps produserer her de største spermiene man kjenner i dyreriket (bare slått av bananfluer): nøttemuslingkreps Cyclocypris ovum har for eksempel spermier som er 6 ganger så lange som de selv, og fossile spermier av ostracoder har vært opptil 1 cm lange (Matzke-Karasz 2006; Matzke-Karasz m.fl. 2014)! Med slike monstrøse kjønnsceller blir det ikke plass til svært mange ad gangen, og for å hjelpe dem inn i hunnene har hannene utviklet et separat organ, Zenkers organ, som er en spiralisert pumpe som besørger overføringen av spermier (se bilde nedenfor).
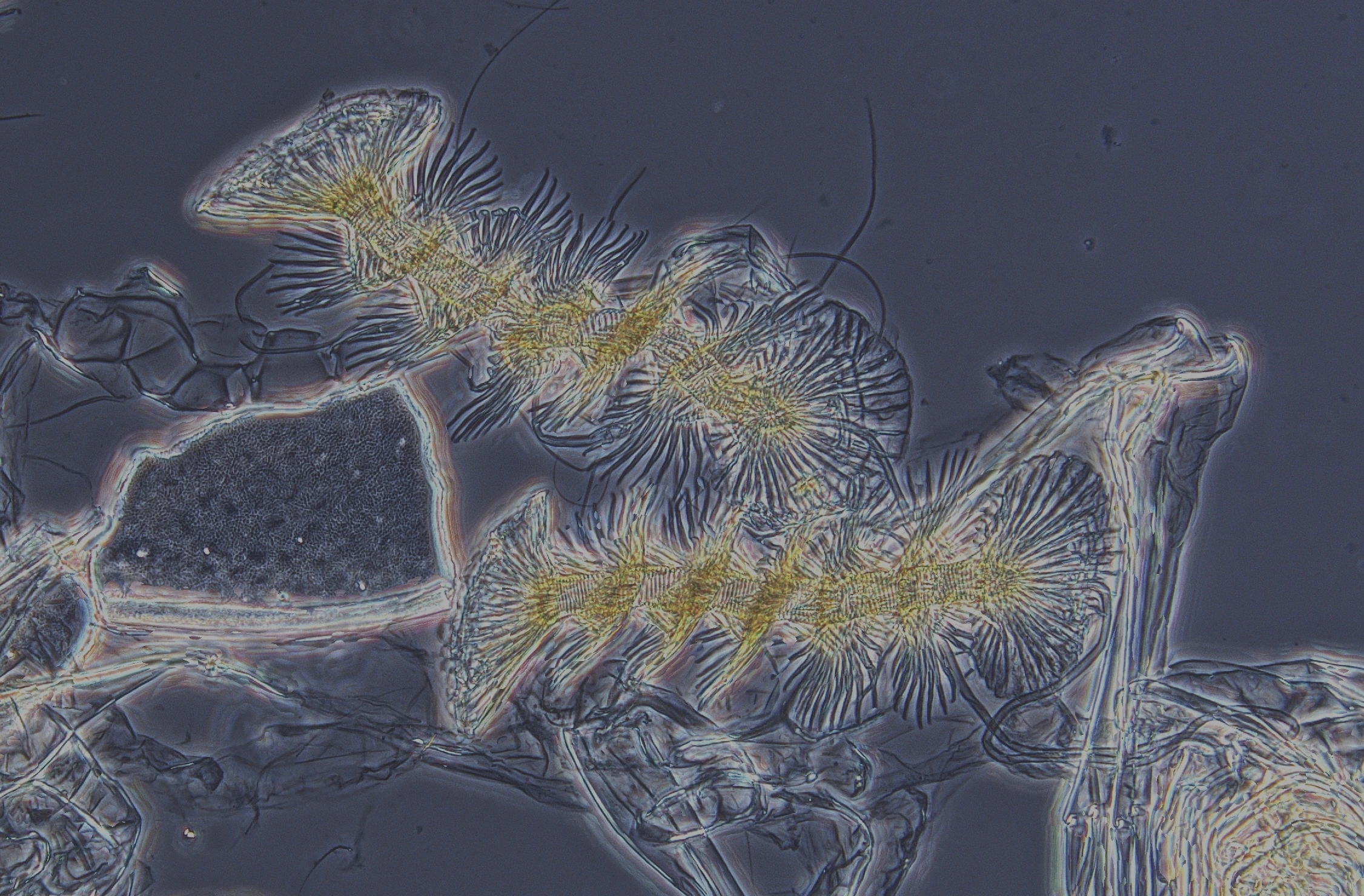
Zenkers organ, en spiralisert pumpe som besørger overføringen av spermier fra hannen til hunnen.
Observasjoner under lupen
For en systematisk og skrittvis identifikasjon anbefales det å gå frem på følgende måte:
Under lupen måler man skall-lengden. For enkelte grupper, blant annet Candonidene (som er den gruppen som erfaringsmessig volder størst problemer), er også bredde/lengde-forholdet viktig (slekten Candona har bredde/lengde-forhold > 0,4, de øvrige slektene innen samme familie har < 0,4). Man noterer seg videre farge, mønstring, overflatestrukturer, behåring, samt randen av skallene, og hvorvidt disse har spiner, hår, pigger eller utvekster. Fargen gir ofte viktig informasjon og artstypisk, gjerne i kombinasjon med bestemte mønstre, og her anbefales det å ta en sjekk på ulike faktaark, for å få et inntrykk av mangfoldet. Noen arter har små pigger langs randen i front (piggmuslingkreps Cypris pubera), andre har små vorter foran på sidene (kumuslingkreps Heterocypris incongruens), eller én markert pigg på skallets nedre rand (sotmuslingkreps Notodromas monacha). Enkelte har speilblanke skalloverflater (Cypriakreps Cypria opthalmica) eller særskilte mønstre eller strukturer (Cypria exculpta). Også gruppen av ovale eller runde former midt på skallet, som er muskelfestene for lukking av skallet, kan gi viktig informasjon (muskelfestene er godt synlige på bildene av stormuslingkreps Heterocypris reptans og på nettmuslingkreps Brandleystrandesia reticulata). Skallets sideprofil er et annet viktig kjennetegn, og etter litt øvelse vil man raskt få blikk for de artstypiske proporsjonene. Særlig viktig er det å legge merke til hvordan rygglinjen går fra nedre kant foran, oppover, bakover og ned i bakkanten. Hvordan krummer linjen seg oppover, hvor bratt stiger den, hvor når den høyeste punkt? Enkelte arter har høyeste punkt foran midten av ryggen (for eksempel nettmuslingkreps Bradleystrandesia reticulata), andre midt på ryggen (bekkemuslingkreps Potamocypris sp.), mens andre har høyeste punkt lenger bak (strihårmuslingkreps Pseudocandona rostrata). Enkelte arter har en svært langstrakt sideprofil (sadelmuslingkreps Dolerocypris fasciata), mens andre er korte og butte (nøttemuslingkreps Cyclocypris ovum).
Når man vipper dem over på «høykant» og ser individene fra ryggsiden (ovenfra) kommer nye viktige trekk til syne. Legg merke til form og bredde på omrisset. Hos noen arter løper sideflatene parallelt, andre har jevnt ovale omriss, mens andre igjen er mer dråpeformet og tilspisset i front. Det er også viktig å se om en av skalldelene overlapper den andre i front eller bak, og om overlappet i så fall er på høyre eller venstre skall. Hos mange arter møtes de to skalldelene i front på en karakteristisk måte – butt mot hverandre (myrmuslingkreps Paracandona euplectella), mer gradvis, eller med utsving som gir avslutningen i front et leppelignende utseende (klomuslingkreps Eucypris pigra). Høyre og venstre skall kan også møtes asymmetrisk, som hos strihårsmuslingkreps (Pseudocandona rostrata), der høyre skall i front har en distinkt konkav innsving, mens venstre svinger jevnt inn mot spissen.
Å tegne skallprofilene kan være en god måte å trene opp blikket for artsspesifikke ulikheter på (bruk en godt spisset myk gråblyant, og ha viskelær klart!). Mange av faktaarkenes fotografier viser omrissene for de ulike artene.
Man vil kunne bestemme mange av artene allerede gjennom slike undersøkelser. Men i motsatt fall fortsetter neste trinn med preparerte individer under mikroskopet.
Preparering og mikroskopiske kjennetegn
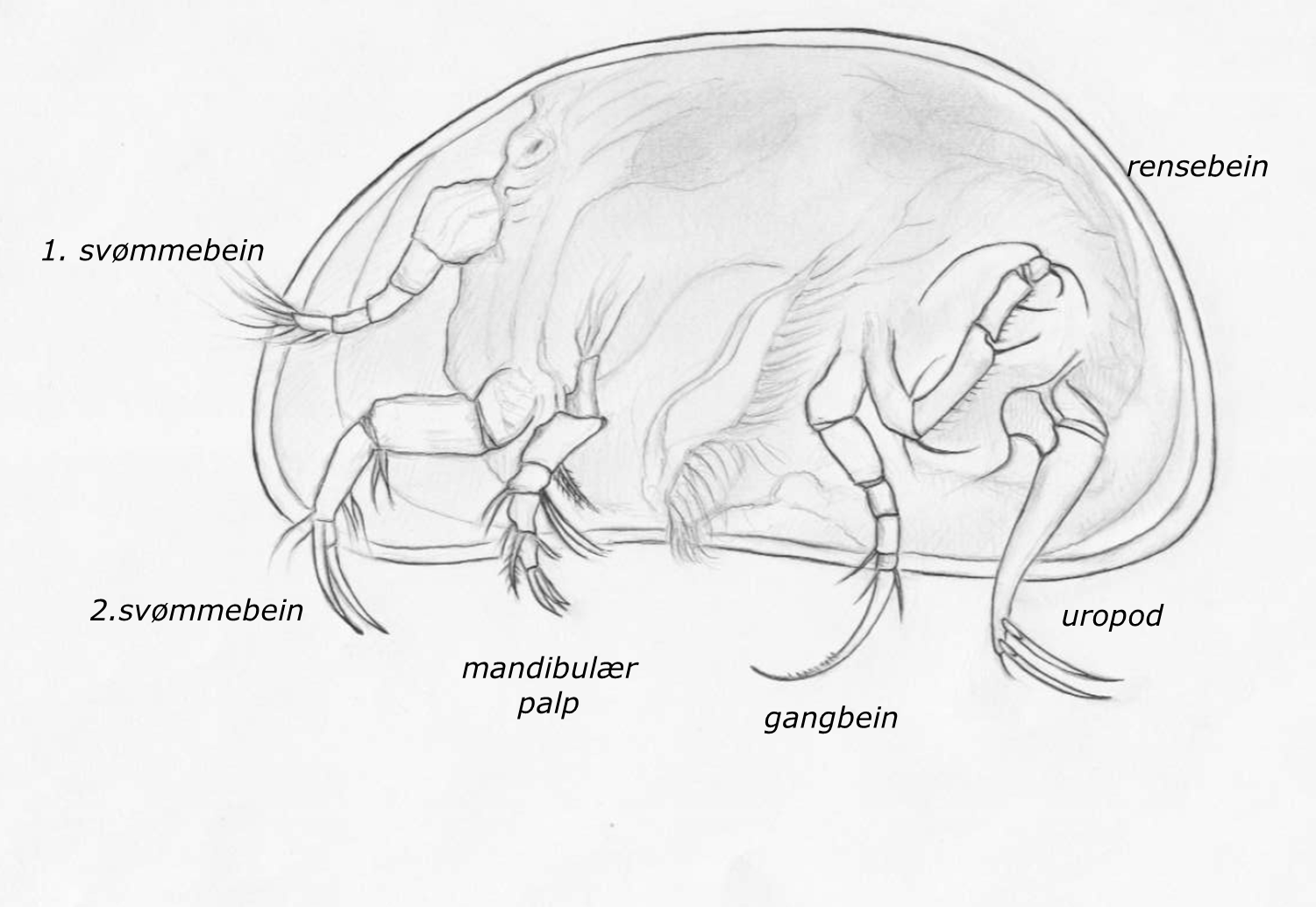
Figur 1
Viktige anatomiske trekk for artsbestemmelse av muslingkreps, her hvitmuslingkreps Candona candida. Alle de markerte strukturene forekommer parvis, plassert på hver side av abdomen. Se teksten for videre detaljer.
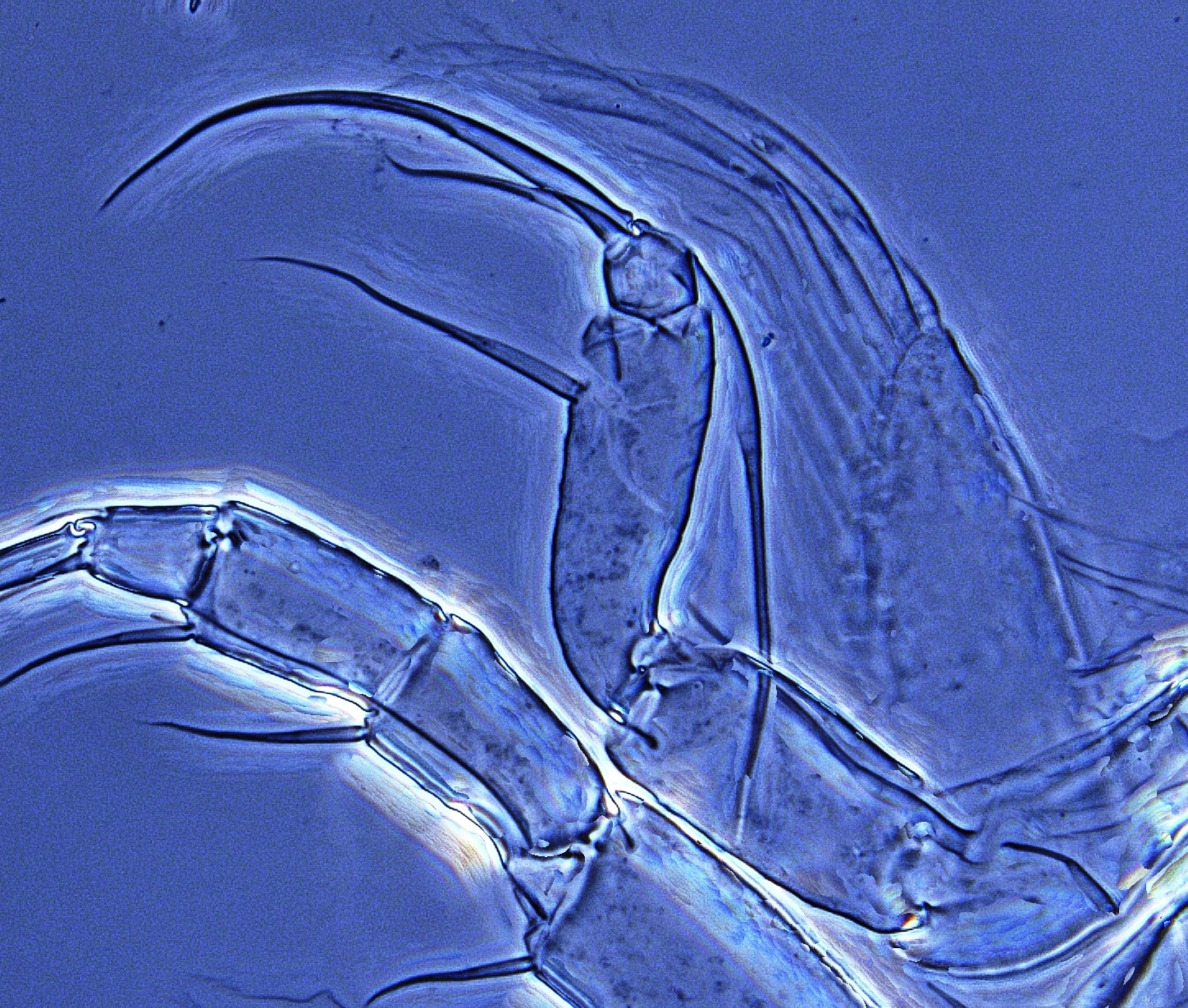
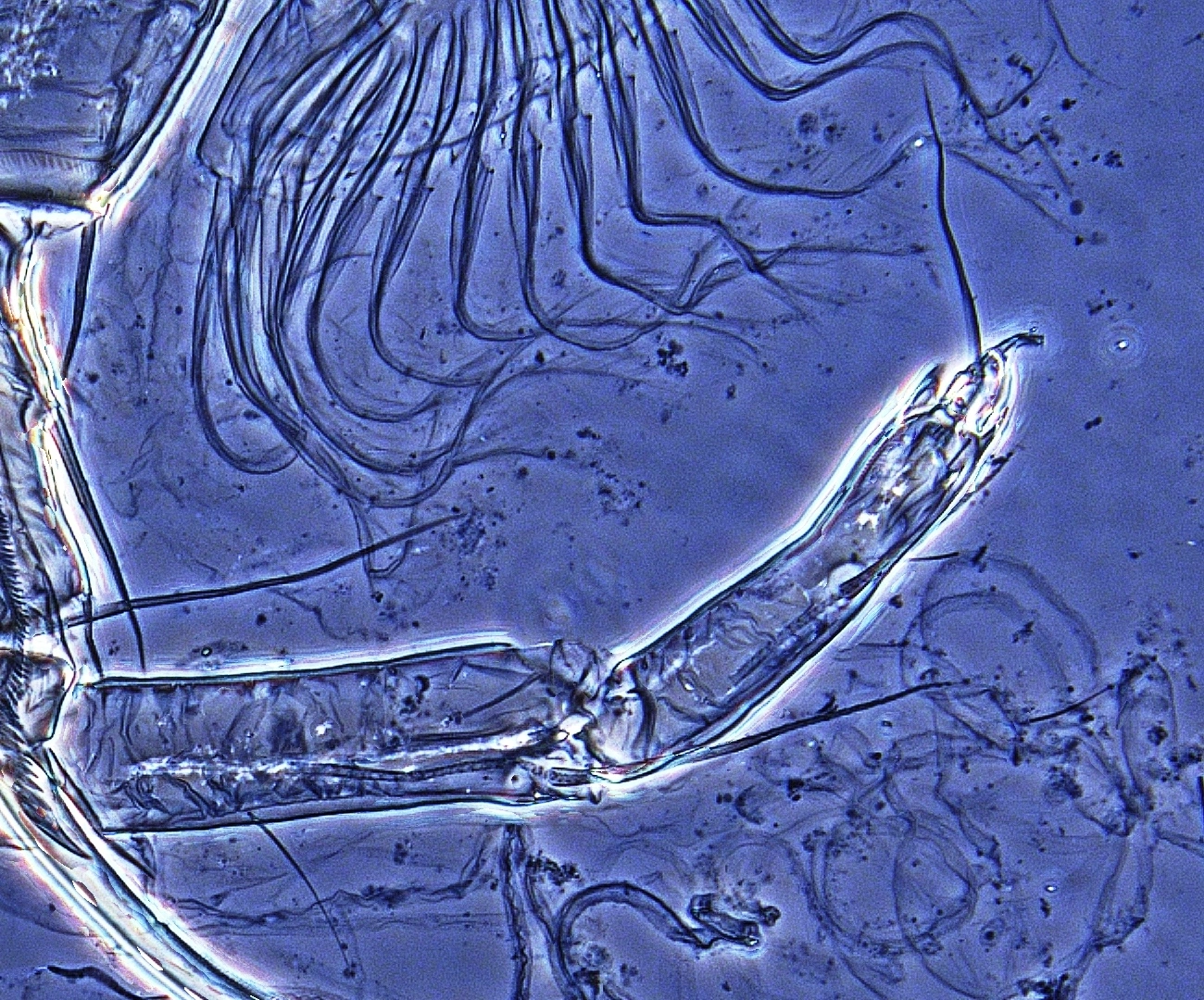
Figur 2
Ytterste del av rensebeina er et viktig kjennetegn for bestemmelse av muslingkreps. Her er de to ulike grunnformene - øverst hvitmuslingkreps (Candona candida), nederst piggmuslingkreps (Cypris pubera).
Det kan lyde vanskelig å preparere muslingkreps for mikroskopering og systematisk bestemmelse, men det er enklere enn man tror, og med litt øvelse går det kjapt. Verktøy som kan komme til nytte er: En spiss pipette, KOH, etanol, eppendorf-rør, fikseringsmedium (glyserol eller euparal), tynne prepareringsnåler, objektglass/dekkglass og små petriskåler. Gode prepareringsnåler kan lett lages selv: Ta ut innmaten av en bic-kulepenn og varm opp enden over en flamme. Når den er bløt skyver man inn en tynn insektnål i plastikken, og lar det stivne.
Med to slike prepareringsnåler har man det man trenger for å manøvrere individene under lupen, og for å åpne skallet når man skal undersøke bløtdeler. De fleste foretrekker å ha muslingkrepsene i en petriskål med etanol mens de undersøkes under lupen. Namiotko m.fl (2011) gir for øvrig en introduksjon til hvordan man kan gå frem for preparering av muslingkreps for mikroskopi.
Man kan gå frem på følgende måte: 4-6 dyr legges i 1 ml KOH og varmes opp til +70-80 °C i 30-60 min (gjerne i en varmeblokk). Etterpå legges de over i vann og skylles forsiktig, men grundig. Med prepareringsnålene kan skallene åpnes noe for å sikre at KOH vaskes godt ut. Dyrene skylles deretter noen minutter i etanol og legges på objektglass i en dråpe glycerol eller euparal. Nå er bare de mineraliserte delene tilbake, og organenes finstrukturer er lett synlige i gjennomlysning. Før dekkglass has på kan de indre delene pirkes ut av skallene, slik at enkelthetene blir noe lettere å se. Fordi dette ikke alltid lykkes kan det være smart å gjøre bruk av 4-6 individer.
Som preparat vil viktige organstrukturer være mye lettere å identifisere. Strukturene med størst relevans for taksonomi er fremhevet på figur 1, og omtales nærmere nedenfor.
På undersiden er det fire par med bein-liknende strukturer, som fra forfra og bakover er: 1. og 2. par svømmebein, eventuelt med hvert sitt knippe av korte eller lange svømmehår (familien Candonidae er kjennetegnet ved at de mangler svømmehårene, og de er derfor helt bentiske), gangbeina, som ender i en klo, rensebeina, der enkelthår og leddenes utforming er viktige kjennetegn for mange arter, og bakerst de to uropodene, som også har klør. Hos sistnevnte er også omrisset av den såkalte kjønnsfolden (eng. «the genital lobe») et viktig kjennetegn (se for eksempel bilde for hvitmuslingkreps Candona candida). Mellom 2.svømmebein og gangbeina sitter den mandibulære palpen, som er bygget opp av fire segmenter, og er bøyd bakover. Det kan trenge litt øvelse å finne denne i mikroskopet, men særlig hos de store slektene av Candonidae (Fabaeformiscandona, Pseudocandona og Candona) er antall og plassering av ulike hår på dette organet viktige artsspesifikke kjennetegn. Det gis en mer spesifikk beskrivelse av de ulike organene nedenfor.
Skall (Carapace): Skallet er todelt i en høyre og venstre del (sett fra ryggsiden) og omgir dyret som et muslingskall. Fylogenetisk er det avledet av ryggskjoldet man ser hos krabber og kreps, men det har forandret form og funksjon. De to delene er bare løst festet sammen dorsalt, med en låsemekanisme som gjør at dyret kan lukke seg helt inne. Dette kan være funksjonelt både om miljøet blir for krevende, eller overfor andre farer. Enkelte arter, som nøttemuslingkreps, kan klype seg fast til andre dyr ned skallet, og forflyttes over større avstander (se faktaark).
Skallformen er ofte artsspesifikk, og både farge, form og overflatestrukturer utgjør en viktig hjelp ved artsbestemmelsene. Overflaten kan være glatt og perlemorslignende hvit, som hos hvitmuslingkreps Candona candida, men den kan også være ornamentert med små ujevnheter (myrmuslingkreps Paracandona euplectella), eller være utstyrt med hår (piggmuslingkreps Cypris pubera). Nærtstående arter som ellers kan være svært like, kan i visse tilfeller skjelnes på skalls overflate. De to artene av slekten Cypria (C. opthalmica og C. exculpta) er ganske like, men førstnevnte har speilblankt skall, mens overflaten hos sistnevnte er besatt med et fint mønster av parallelle rilleformete striper. Hos flere arter har skall også tydelige farger. Hos enkelte arter (hvitmuslingkreps, nøttemuslingkreps) er fargen et sikkert kjennetegn. Men unntak finnes. Nettmuslingkreps Bradleystrandesia reticulata er gjerne akvamaringrønn, men man kan også finne lysgrønne, blålige eller blekgrå individer. Fargene kan også være vakkert marmorert, eller ha striper eller «sadel». Stripemuslingkreps Cypridopsis vidua har fire distinkte tverrstriper i grønt, mens sadelmuslingkreps Dolerocypris fasciata og Bradleystrandesia fuscata har én bred tverrgående sadel.
Uropodene er et par langstrakte, muskuløse beinstrukturer bakerst. De ender ytterst i ett par med såkalte furca-klør, den ene kloen gjerne lenger enn den andre. Uropodenes form og lengde er viktige kjennetegn, samt ligamentene de er festet til. Mange arter har ett markert hår på undersiden av uropoden, og både lengden og plasseringen av dette kan være viktig. Rett ovenfor og nær uropodens feste har mange arter en karakteristisk fold, kalt kjønnsfolden (på illustrasjonen markert som en liten bueformet utposning; se faktaark hvitmuslingkreps for fotografisk eksempel). Uropodens utforming og kjønnsfolden er særlig viktige kjennetegn for den artsrike underfamilien Candonidae (for eksempel slektene Candona, Pseudocandona, Fabaeformiscandona). Unntaksvis mangler uropoden, slik som hos bekkemuslingkreps (Potamocypris sp.), der den er omdannet til en liten avsmalnende vevsstruktur som ender i ett langt hår.
Gangbeinet er plassert nært uropoden, og er tett koplet til rensebeinet. For å finne det siste i preparatet kan det være klokt først å lete frem gangbeinet, som vanligvis er iøynefallende med den lange, krummede enkeltkloen. Gangbeinet kan være bygget opp av fire eller fem ledd, og er et viktig kjennetegn for familien Cyprididae. Forskjellen beror egentlig på at det nest ytterste leddet er udelt (slik som hos for eksempel piggmuslingkreps Cypris pubera) eller delt (slik som for eksempel hos sadelmuslingkreps Dolerocypris fasciata eller stripemuslingkreps Cypridopsis vidua).
Rensebeinet er den første viktige mikrostrukturen man bør lære seg, og den er det første trekket man sjekker under mikroskopet. Det ytre enden avgjør den videre bestemmelsen. Enten er denne delen utformet som et separat ledd, besatt med tre hår av ulik lengde (figur 2, øverst), eller det mangler, og enden er formet som et særegent, klypeformet renseorgan (figur 2, nederst). Den første versjonen er felles for alle Candonider og Cyclocypris sp., mens de øvrige har «klypeløsningen» felles. I førstnevnte tilfelle, altså der enden har form av et separat lite ledd, er dette besatt med tre hår, og lengde og form på disse gir viktig informasjon for artsbestemmelsen. Og for Candonider er også andre deler av rensebeinet viktig for artsbestemmelsen. Alle arter av slekten Pseudocandona har for eksempel tre hår på basalleddet (de øvrige har to). Slekten Cryptocandona (to arter i Norge, som ikke er videre omtalt her) kjennetegnes av at det nest ytterste leddet på rensebeinet er delt, og hver del distalt bærer ett hår.
Den mandibulære palpen kan i begynnelsen være vanskelig å finne, men er viktig for artsbestemmelse av en del viktige artsgrupper. Hos Candonider er den krummet, og på krummingens innside sitter et antall lange parallellstilte hår, der antallet (vanligvis 3, 4 eller 5) er viktige for den videre bestemmelsen. hos slekten Potamocypris sp. er det distale leddet på den mandibulære palpen bredt og spatelformet utvidet.
2. svømmebein gir også viktig informasjon. Det er leddet, og ender i et knippe klør. Disse er gjerne like lange, men hos enkelte arter er en av klørne markert kortere. Videre er det midtre leddet gjerne besatt med et knippe myke hår, som styrker svømmeevnen. Hos enkelte arter er de svært mye lenger enn klørne, og slike arter er gjerne gode svømmere. Hos andre danner hårene bare en kort bust, for eksempel hos Potamocypris pallida. Candonider mangler helt disse hårene på 2.svømmebein, og er derfor helt bunnlevende.
Referanser
Alm G (1914). Beschreibung einiger neuen Ostracoden aus Schweden. Zoologische Anzeiger 43:468-475.
Alm G, (1915). Monographie der schwedischen Süsswasser-Ostracoden nebst systematischen
Besprechungen der Tribus Podocopa. Zoologiska Bidrag från Uppsala, 4: 1-248.
Langeland A (1972). Hjelpetabell for bestemmelse av norske ferskvanns-ostracoda. NIVA notat.
Matzke-Karasz R (2006). Giant spermatozoon coiled in small egg: Fertilization mechanisms and their implications for evolutionary studies on ostracoda (crustacea). Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 304B/2:129-149. DOI: 10.1002/jez.b.21031
Matzke-Karasz, R., Neil, J.V., Smith, R.J., Symonova, R., Morkovsky, L., Archer, M., Hand, S.J., Cloetens, P., Tafforeau P. 2014. Subcellular preservation in giant ostracod sperm from an early Miocene cave deposit in Australia. Proc. R. Soc. B 20140394. http://dx.doi.org/10.1098/rspb.2014.0394
Meish C (2000). Freshwater Ostracoda of Western and Central Europe. Süsswasserfauna von Mitteleuropa 8/3. Spektrum Akademischer Verlag, Heidelberg.
Namiotko T, Danielpol DL og Baltanas A (2011). Soft body morphology, dissection and slide-preparation of Ostracoda: a primer. Joannea Geol. Palaeont. 11:327-343.
Lenker
International Research Group on Ostracoda: http://www.irgo.uni-koeln.de/
North American Non-Marine Ostracode Database Project: http://www.personal.kent.edu/~alisonjs/nanode/index.htm
Ostracod Research at the Lake Biwa Museum, Japan. Key to Genera of Japan: http://www.lbm.go.jp/smith/keygenera.html