Humler
Bombus

Dronning av kløverhumle Bombus distinguendus på rødkløver, Rinnleiret i Nord-Trøndelag, juli 2013.
Humlene tilhører de mest iøynefallende insektene i vår del av verden. De utgjør viktige komponenter i økosystemer gjennom sin avgjørende rolle som bestøvere (pollinatorer) for en rekke planter. På våre breddegrader er humlene både tallrike og vidt utbredt i mange naturtyper fra kyst til fjell. Av verdens mer enn 250 humlearter er hele 35 påvist i Norge.
- Innhold
- Arter i Norge
- Systematikk
- Sosiale humler
- Jordhumler
- Gjøkhumler
- Habitatvalg og blomsterpreferanse
- Humler i fjellet
- Revirhevding, flukt og paring
- Konkurranse
- Fiender og sykdommer
- Genetikk
- Endringer i kulturlandskapet
- Klimaendringer
- Fremmede planter
- Giftige planter og biltrafikk
- Humler som modeller
- Kartlegging av humler i Norge
- Bestemmelse av humler
- Kroppsbygning
- Praktiske tips
På tilsvarende måte som for fugler og sommerfugler, er humler populære både blant naturinteresserte og folk flest. Det skyldes trolig at de er forholdsvis store, fargerike og dagaktive, samt den økende bevissthet om at de gjør en nyttig jobb for oss mennesker og er viktige indikatorer for naturens miljøtilstand. Mange av humleartene har karakteristiske fargemønster som gjør at de lett kan artsbestemmes. Humler er fredelige insekter, som bare ytterst sjelden stikker hvis de for eksempel kommer i klem. Det sies at humlene brenner i stedet for å stikke, men det skyldes kanskje at stikkene fra giftbrodden gir en brennende smerte. Det er bare dronninger og arbeidere som har giftbrodd.
Arter i Norge
På verdensbasis finnes det i overkant av 250 humlearter, mens det hittil er påvist 35 arter i Norge. Humlene dominerer på den nordlige halvkule, og det er ikke mange organismegrupper der nesten 14 % av verdens arter finnes hos oss. Mange humlearter har gått sterkt tilbake både i Europa og Nord-Amerika som følge av omfattende omlegging i landbruket de siste 100 år. Også klimaendringer, pesticider og introduksjon av fremmede humlearter, ser ut til å ha slått tilbake mange arter (Goulson 2010, Williams & Osborne 2009). Likevel har Norge et rikt mangfold av humler, og de fleste artene som finnes hos oss har livskraftige og store bestander.
Noen av våre aller vanligste arter, som både har vid utbredelse og store bestander i mange naturtyper, er: trehumle, lys jordhumle, mørk jordhumle, hagehumle, lundhumle, lynghumle, steinhumle, åkerhumle, markhumle, jordgjøkhumle og markgjøkhumle. De mest sjeldne humleartene i Norge er kløverhumle, slåttehumle og lundgjøkhumle. Noen arter er begrenset utbredt til kystområder, slik som kysthumle og delvis kragejordhumle. Fjellet har en egen humlefauna med fjellhumle, alpehumle, polarhumle, tundrahumle, berghumle, lapphumle og lynggjøkhumle. Arter som tyvhumle, taigahumle, kilejordhumle og barskoghumle er mest vanlig i høyereliggende skogsområder. Videre har vi noen arter som hovedsakelig påtreffes i lavlandet på Østlandet slik som gresshumle, enghumle, bakkehumle og slåttehumle.
Vi forventer at ytterligere to humlearter vil bli oppdaget i Norge i løpet av de nærmeste årene. Den ene er hagegjøkhumle Bombus barbutellus som er sosialparasitt på hagehumle Bombus hortorum og er funnet i Sverige ikke så langt fra grensa mot Norge i Hedmark og Østfold. Hagehumla ser ut til å ha blitt mer vanlig igjen i Norge de siste årene blant annet som følge av økningen i den fremmede planten kjempespringfrø Impatiens glandulifera. Det er derfor ikke utenkelig at hagegjøkhumla snart vil dukke opp hvis den ikke allerede er her.
Den andre arten er praktgjøkhumle Bombus vestalis som er sosialparasitt på mørk jordhumle. Siden mørk jordhumle er på rask spredning i Skandinavia, er det å forvente at praktgjøkhumle vil følge etter. Praktgjøkhumle ble i Sverige først oppdaget i 1973 og har siden spredt seg i de sørligste delene hvor den nå er den vanligste gjøkhumlearten flere steder.

Blåknapp er en populær plante blant humlene når sommeren er på hell og de fleste andre plantene er avblomstret, Aurskog-Høland i Akershus.
Systematikk
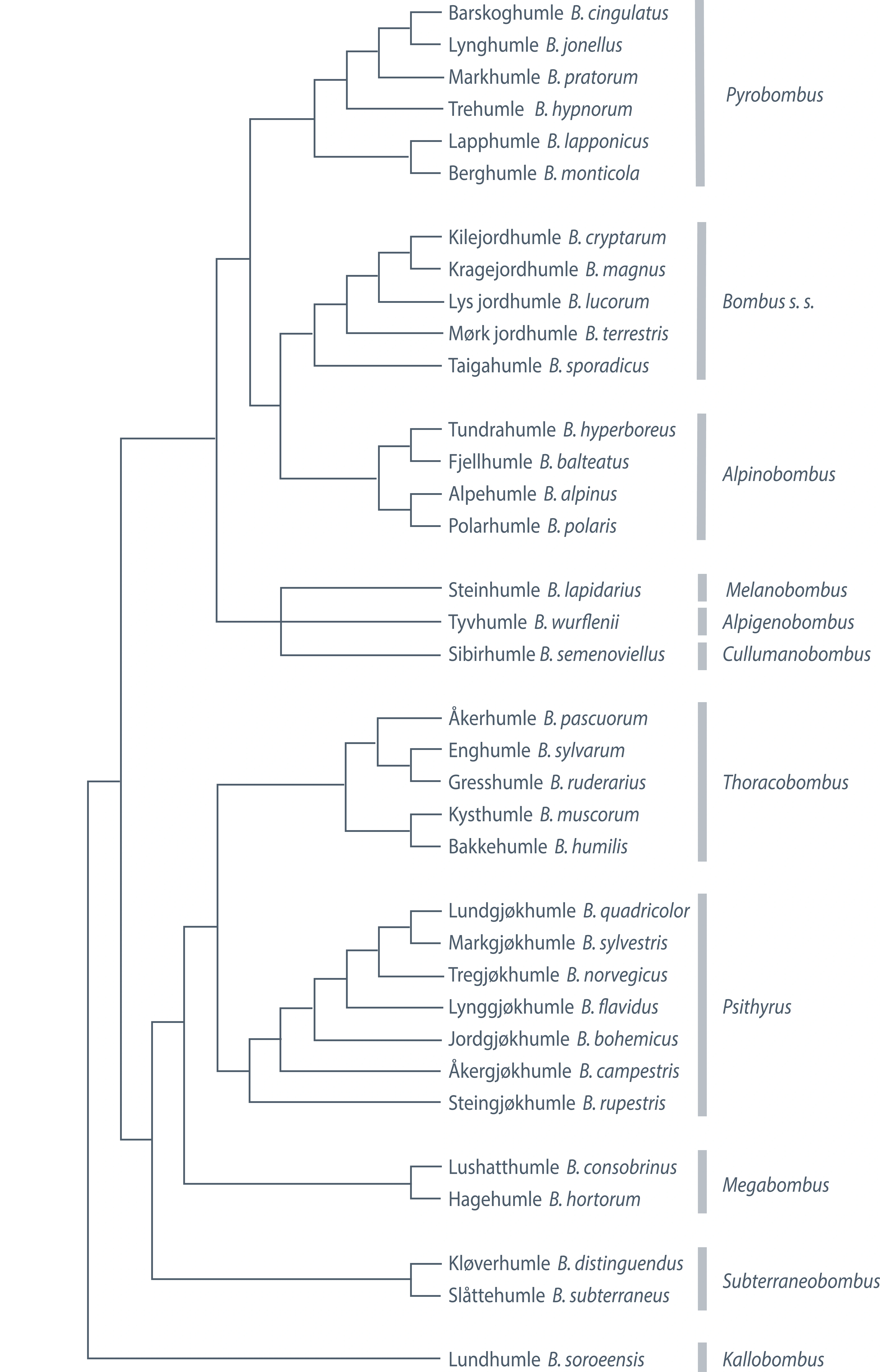
Slektskapet mellom de 35 humleartene som er påvist i Norge og deres tilhørighet til ulike underslekter. Omarbeidet etter Cameron et al. (2007).
Humlenes fylogenetiske tre er basert på DNA-analyser av fem gener (Cameron et al. 2007). Denne analysen viste at gjøkhumlene ikke kunne skilles ut som en egen slekt, Psithyrus. De blir nå behandlet som en av verdens 15 underslekter av humler, hvorav 11 er representert i Norge. Kunnskap om artenes slektskap er nyttig for å forutsi trekk ved deres biologi når dette er kjent hos nærstående arter. Langtungete humler hører hjemme i underslektene Thoracobombus, Megabombus og Subterraneobombus. Dette er samtidig humlene som tilhører "lommebyggerne" (pocket makers), som inngår i den mest primitive form for yngelpleie (se nærmere omtale under sosiale humler). Det er grunn til å anta at alle artene i disse underslektene er lommebyggere, selv om yngelpleie hos alle aktuelle arter ennå ikke er beskrevet.
Sosiale humler
Av de 35 humleartene som er påvist i Norge, er 28 sosiale. At de er sosiale betyr at de lever sammen og danner samfunn der ulike individer deler på arbeidsoppgavene som inkluderer blant annet bolbygging, ruging, yngelpleie, vakthold og sanking av føde. Humler i vår del av verden har ettårige samfunn som består av kastene dronninger, arbeidere og hanner. Dronninger er store og fruktbare hunner som parer seg på høsten og etablerer nye bol neste vår/forsommer. Arbeiderne er mindre hunner som ikke pares, men som i noen tilfeller kan legge egg som utvikles til hanner. Det er ikke kjent hvor stor andel av hannene som stammer fra arbeidere eller dronninger. Hannene er haploide (har et enkelt sett med kromosomer), mens dronningen og arbeiderne er diploide (har dobbelt sett med kromosomer). Arbeiderne har forskjellige arbeidsoppgaver. Noen er utearbeidere som samler inn pollen og nektar, mens andre ruger egg og steller larver, og er med på å holde bolet i orden. Arbeidsoppgavene glir ofte over i hverandre og kan variere i løpet av en humles liv.
Det er bare den befruktete dronningen som overvintrer. Både gamle dronninger, arbeidere og hanner dør før vinteren. Om våren eller forsommeren anlegger dronningen et bol etter at hun har våknet opp av vinterdvalen, gjerne inne i et gammelt bol av smågnagere på eller under bakken. Noen arter kan også anlegge bol i et fuglereir i en reirkasse eller et hult tre. Dronningen bygger en liten krukke av tynne plater av voks som hun skiller ut fra undersiden av bakkroppen. Krukka fylles med humlehonning som et reservelager av proviant, som blant annet kommer godt med under ruging og på dager med dårlige værforhold. I tillegg bygger hun yngelceller av voks hvor eggene legges på toppen av en klump med pollen blandet med honning, og det hele dekkes med voks (se bilde av honningkrukke og voksklump). Dronningen ligger og ruger på eggene til de klekker. Ved å viberere med brystmusklene, produserer hun varme. Kroppstemperaturen hennes kan komme opp i 37–39 ⁰C. Larvene vokser hurtig og gjennomgår fire stadier før de spinner en silkekokong og forpupper seg. Det tar 4–5 uker fra egg til voksen humle, avhengig av temperatur og fødetilgang. En vanlig kullstørrelse er åtte. Det første kullet av humlearbeidere på forsommeren består av små individer.
Det er to hovedtyper av yngelpleie hos humler. Den ene er den såkalte ”pollen storers” (pollenlagrere) som lagrer pollen direkte i tomme kokonger. Deres larver blir foret av dronningen eller arbeidere ved at en blanding av pollen og honning blir gulpet opp til dem. Dette blir ansett å være den mest avanserte form for yngelpleie. De fleste av våre sosiale humlearter som jordhumlene, lundhumle, steinhumle, tyvhumle, trehumle, lynghumle, markhumle, fjellhumle, alpehumle, lapphumle og berghumle tilhører denne kategorien. Disse artene regnes samtidig som korttungete, selv om det i praksis er små forskjeller i tungelengde hos noen korttungete og langtungete arter.
Den andre formen for yngelpleie kalles ”pocket makers” (lommebyggere). Hos disse artene oppbevares pollen i egne vokslommer som bygges på utsiden av yngelcellene. Larvene eter selv av pollenlageret. Av våre humler tilhører hagehumle, lushatthumle, kløverhumle, slåttehumle, bakkehumle, kysthumle, åkerhumle, gresshumle og enghumle denne kategorien. Dette er de såkalte langtungete humleartene, og samtidig de mest primitive i et evolusjonært perspektiv.

Honningkrukke og voksklump
Begynnende bol hos åkerhumle Bombus pascuorum, Trondheim i Sør-Trøndelag. Dronningen starter samfunnet ved å bygge en honningkrukke som fylles med humlehonning. Deretter legges de første eggene, ca. 10, i en klump av pollen og nektar som dekkes av voks. Dronningen ruger på eggene ved å avgi varme gjennom kroppen fra vibrasjoner i flygemuskulaturen. Eggene klekkes etter ca. fire-fem dager.
Jordhumler
Jordhumlene nevnes ofte som en enhetlig gruppe av arter og tilhører våre aller vanligste humler. Fem arter inkluderes i denne gruppen i Norge: lys jordhumle, kilejordhumle, kragejordhumle, mørk jordhumle og taigahumle. Alle disse, unntatt taigahumle, kan være svært krevende å skille fra hverandre. For sikker artsidentifisering av vanskelige individer, kreves genetiske analyser (DNA-strekkoding av CO1-genet i mitokondriene) (Williams et al. 2012).
Mørk jordhumle er på en måte i en særstilling siden denne avles opp og brukes i veksthus som pollinator av nyttevekster, særlig tomater. Arten finnes naturlig i Norge, men ble ikke påvist her før på 1950-tallet. Den har siden ekspandert voldsomt og er funnet nord til Narvik. Nå er den ofte den vanligste humlearten i bynære strøk i Sør-Norge. Ekspansjonen av mørk jordhumle har trolig sammenheng med både rømminger fra veksthus og et gunstigere klima for arten hos oss. Bruk av mørk jordhumle i veksthus i Norge er omdiskutert på grunn av mulige negative konsekvenser knyttet til rømming og etablering på friland. Dette fordi den antas å konkurrere med hjemmehørende arter om blomsterressurser og bolplasser, samt at importerte humler kan bringe med seg smittsomme sykdommer. Man er også redd for at importerte humler kan bidra til genetisk endring i hjemmehørende bestander av mørk jordhumle (Gjershaug & Ødegaard 2012).

Dronning av mørk jordhumle Bombus terrestris på blomstrende lønn, Hovedøya i Oslo.
Gjøkhumler
Vi har sju arter ekte gjøkhumler i Norge. Disse tilhører underslekten Psithyrus som tidligere ble regnet som en egen slekt. De samler ikke nektar og pollen, men fortærer det umiddelbart. Gjøkhumlene har derfor ikke pollenkurver, som de sosiale humlene, men har rundere og matte bakskinnebein med jevnlang behåring. De har noe tykkere kutikula, og hunnene har svært glissen behåring på bakkroppen og kraftig stikkebrodd. Bakkroppsspissen er mer eller mindre nedover- og innoverbøyd. De har videre ganske kraftige kjever som ender skrått slik at nedre endehjørne danner en spiss vinkel. Gjøkhumlene har i tillegg ganske kort tunge og ofte mørkere vinger. De virker ganske trege og har som regel en mer lavfrekvent summetone sammenlignet med sosiale humler.
Gjøkhumlene er sosiale parasitter og mangler arbeidere. De overtar bolet til sosiale humlearter ved at de dreper eller jager vertsdronningen og legger sine egne egg i vertens bol. Hunnene av gjøkhumler våkner ganske seint etter vinterdvalen, for de okkuperer ikke et bol før det har kommet arbeidere der. Vertens arbeidere steller egg og larver, som bare utvikler seg til dronninger (hunner) og hanner.
På våren er løvetann en favorittplante for hunnene. Utpå sommeren klekker store mengder hanner av gjøkhumler, som er lette å oppdage der de sitter dorske på planter med mye nektar og pollen. Rødknapp og blåknapp er populære, men også ulike andre seintblomstrende kurvplanter som tistler, knoppurt, svever og dyller. De nye hunnene kommer også for å ta til seg næring før overvintring, men de er ikke så ofte å se.
Vi har enda en sosialparasittisk humleart; tundrahumle. Den overtar bolet til alpehumle, polarhumle eller fjellhumle, men kan i tillegg av og til også produsere noen få egne arbeidere. Tundrahumle regnes blant de sosiale humlene og tilhører fjellhumlene (underslekten Alpinobombus).

Hanner av gjøkhumler er veldig vanlige å finne på blomster på ettersommeren, og rødknapp er en av favorittene. Bildet viser tregjøkhumle Bombus norvegicus, Oppdal i Sør-Trøndelag.
Habitatvalg og blomsterpreferanse
Humlene finnes så og si overalt i Norge fra kyst til høyfjell, men de ulike artene har til en viss grad delt naturtypene mellom seg. De fleste arter har riktignok preferanse for åpne habitater der det er rikelig med blomster og muligheter for bolplass. Det er derfor få humler i tett skog, men mange arter dukker opp i små lysninger, på hogstflater og i kantsoner. Når selje- og viertrær blomstrer i skogen, er det dessuten et rikt humleliv i trekronene. I det åpne landskapet i lavlandet finnes et rikt humleliv i tilknytning til blomsterrike områder som kystheier, åkerkanter, veikanter, slåtteenger, ruderatområder og ikke minst i hagene våre. Det er enkelt å legge til rette for humler i sin egen hage ved for eksempel å sette av en del av plenen til slåtteeng og å plante humlevennlige planter. Man kan også sette ut humlekasser som bolplasser til humlene.
Humlene er avhengige av å finne blomster med pollen og nektar gjennom hele sesongen. Mange blomster er tilpasset å bli bestøvet av humler. Blomstene signaliserer med form, farge og duft at de er rike på pollen og nektar. De har gjerne blå, fiolette eller lyst rødlige farger. Humler er vanligvis ikke så strengt spesialiserte til enkelte blomsterarter siden humlesamfunnets levetid oftest er lengre enn en enkelt plantearts blomstringstid. Enkelte humlearter er mer spesialiserte enn andre. Det beste eksemplet på en spesialisert art er lushatthumla, som i hovedsak oppsøker blomster av tyrihjelm (lushatt). Den har en ekstremt lang tunge som når helt inn til forekomsten av nektar. Tyrihjelmen har lang blomstringstid og kan dermed forsyne lushatthumla med næring lenge nok til at den kan fullføre sin livssyklus. Utbredelsen av lushatthumla samsvarer svært godt med utbredelsen av tyrihjelm. De spesialiserte humleartene kjennetegnes ofte ved at de har en forholdsvis kort sesong på grunn av at de prefererte fødeplantene har en begrenset blomstringstid.
Gode humleplanter omfatter særlig planter i erteblomstfamilien som kløver og vikker. Men også flere arter i lyngfamilien, leppeblomstfamilien, korgplantefamilien og rosefamilien er gode humleplanter. Mange av de langtungete humleartene er avhengige av planter i erteblomstfamilien som har et særlig proteinrikt pollen, og som i tillegg har dype og smale kronrør godt tilpasset humler med lang tunge. Lundhumla går mye på blomster av blåklokke, men bruker gjerne storkenebb på forsommeren før blåklokkas blomstringstid.
Lengden av humlas tunge avgjør hvor dype og trange blomster den kan drikke nektar fra. Innsamling av nektar er mest effektiv når tungelengden passer til lengden på blomstens kronrør. Noen av artene med kort tunge, som tyvhumla og jordhumlene, kan stjele nektar uten å bestøve blomsten. De biter hull i bunnen av kronrøret og drikker nektaren gjennom hullet.

Lushatthumle Bombus consobrinus er spesialist på å hente nektar fra blomster av tyrihjelm (lushatt). Den har en ekstremt lang tunge (nesten på lengde med kroppen) som når helt inn til bunnen av blomsten der nektaren lagres.
Humler i fjellet
Nesten halvparten av Norges landareal består av fjell og vidder. Den som har sansene med seg på fjellvandringer, og leter på de rette stedene, finner fort ut at også fjellet har en rik humlefauna. I slutten av mai eller begynnelsen av juni, når vierbuskene begynner å blomstre, er det lett å oppdage store mengder av humledronninger som samler pollen og nektar. Utover sommeren, når blomstringen er mer spredt, er det vanskeligere å finne de alpine humlene. Og faktisk er det viktigere å bruke ørene enn øynene på humlejakt i fjellet. Det er lett å høre lyden av ei humle i fjellterreng, men til tross for skarpe farger og stor kroppsstørrelse er det ofte vanskelig å lokalisere humlene når det er store arealer og lave planter i omgivelsene. Mange av fjellviddene virker nokså fattige uten særlig mye blomster om sommeren, og da er det heller ikke mange humler å finne. Det gjelder altså å finne blomsteransamlinger i fjellet om man vil studere de alpine humlene.
Fjellet har altså en helt egen humlefauna, men mange av de vanlige lavlandsartene kan også opptre over tregrensa. I lavalpin vegetasjonssone, der det er rikelig med vier og lyngvekster, er det særlig arter innenfor underslekten Pyrobombus som dominerer (særlig berghumle, lapphumle, lynghumle), samt lynggjøkhumle. Arter som lys jordhumle, kilejordhumle, markhumle, tyvhumle, åkerhumle og trehumle kan også følge med til over tregrensa, men er ikke så vanlige der som i lavlandet. Når vi beveger oss oppover i høyden, til mellomalpin vegetasjonssone, er det fjellhumlene (underslekten Alpinobombus) som dominerer. De fire artene i denne underslekten (fjellhumle, alpehumle, polarhumle og tundrahumle) trives best i kalkrike områder i fjellet. Artene har relativt lang tunge og henter derfor nektar helst fra erteplanter. På ekstrem-kalkrike fjellheier er det ofte mye av favorittplantene setermjelt, blåmjelt og reinmjelt.
I gammel litteratur angis ofte 1300 eller 1400 moh. som maksimal høydegrense for fjellhumlene (f.eks. Løken 1973). Hvis en er heldig med været, og går på de rette stedene, er det nå for tiden ikke uvanlig å finne alle fjellhumleartene på 1500 moh. i fjellet i Sør-Norge, og trolig enda høyere i kalkrike, klimatisk gunstig beliggende hellinger og på toppene. Og det er ikke bare at humlene kan hente næring så høyt til fjells. Sommeren 2014 ble det funnet et bol av alpehumle på en fjelltopp hele 1524 moh. på Dovrefjell (F. Ødegaard upubl. data). Vi kjenner ikke til at humlebol er funnet høyere til fjells i Norge. At fjellhumlene anlegger bol på fjelltoppene, er neppe en tilfeldighet. Bortsett fra at de må kunne takle det tøffe klimaet, er det trolig flere fordeler med å bo på toppen. For det første er det tidlig snøfritt på fjellheiene og rabbene, men det kan også ha sammenheng med at humlene samler seg på toppene for næringssøk eller paring. Samtidig kan toppunktet være plassert i optimal avstand fra flest mulige matkilder. I hviken grad fjellhumlene regulært anlegger bol på fjelltoppene gjenstår å se.
Kanskje er det klimaendringer som har ført humlene oppover i høyden, men det er også mulig at folk i liten grad har lett etter humler i høyfjellet før i tiden. Dersom humlene virkelig trekker oppover i høyden, kan dette være et problem der fjellene er lave og humlene ikke har noen steder å flykte når toppen er nådd. Dette er imidlertid neppe et problem særlig mange steder i Norge, men i flere andre land har fjellhumlene havnet på rødlista som følge av mildere klima. På samme måte som for andre fjellinsekter, kan en finne ansamlinger av fjellhumler på fjelltopper. Dette gjelder særlig på varme finværsdager med lite vind utpå sommeren. Hos for eksempel reinbrems antar man at dette er en effektiv strategi for å finne en paringspartner. Om våren er det heller ikke uvanlig å finne de fire fjellhumleartene under tregrensa. Men etterhvert som snøen trekker seg tilbake og blomstringen starter høyere oppover i fjellet utover sommeren, trekker disse humlene oppover i høyden.

Fjellheier med kalkrik berggrunn i mellomalpin sone er perfekte habitater for fjellhumler (Alpinobombus). Bildet er fra toppen av Finnshøa (1440 moh.), Dovrefjell (Oppdal, Sør-Trøndelag).
Revirhevding, flukt og paring
Hanner av noen humlearter, inkludert slåttehumle og gresshumle, oppsøker unge dronninger og parer seg med dem når de kommer ut av bolet. De kan skille dronningene fra arbeidere på lukten. Det er kjent at hanner kan fly mellom flere bol som ligger i nærheten av hverandre.
Den vanligste måten å finne make på for humler, inkludert gjøkhumler, er at hanner patruljerer faste ruter, alltid i samme retning. Dette ble beskrevet av Darwin allerede i 1886. Hannene markerer ulike steder langs ruta med feromoner. Feromonene er forskjellige hos ulike humlearter, noe som i en del tilfeller også kan registreres med menneskets nese. Disse kjemiske forbindelsene er også brukt i taksonomiske studier. Dette var av stor betydning da lapphumle i Skandinavia ble skilt i to arter, lapphumle og berghumle. Hanner av disse to artene har så forskjellig lukt at vi kan kjenne det.
Ulike humlearter markerer i bestemte høyder over bakken. Hagehumle og markgjøkhumle markerer mindre enn en meter over bakken, mens steinhumle, lys jordhumle og mørk jordhumle flyr i tretopphøyde på opptil 17 meters høyde. Paring hos humler blir observert svært sjelden. Dette er forståelig ettersom det ofte skjer i tretoppene. Unntaksvis kan det observeres på blomster, på trestammer eller vegger nærmere bakken. I fjellet er det som nevnt observert at humlehanner ofte samler seg på fjelltoppene. Dette kan være en strategi for å finne en paringspartner på samme måte som hos mange andre insekter i fjellet.
Dronninger flyr ofte over store avstander på jakt etter ny bolplass om våren eller i søk etter blomster. En kan ofte se at de krysser store kornåkrer eller vann. Hos mørk jordhumle har en målt flygeavstander på opptil 9,8 km. De kan fly med en hastighet på mer enn 15 km i timen. Åkerhumle hører til de arter som søker føde nærmere bolet, med lengste registrerte avstand på 800 meter. Arter med store kolonier har en tendens til å fly lengre enn arter med små kolonier, da de har et større næringsbehov. Fjellhumlene (Alpinobombus) kan også åpenbart fly over lange avstander da de av og til kan ses i målrettet flukt i høy fart og høyt oppe i lufta.

Paring hos jordhumler Bombus sp. på bergknapp (Sedum sp.), Titran i Frøya, Sør-Trøndelag, 1. oktober 2010. Dette er høyst trolig kilejordhumle Bombus cryptarum, men eksemplarene ble ikke samlet inn og kan derfor ikke artsbestemmes sikkert.
Konkurranse
Mens klimatiske forhold og geografisk beliggenhet bestemmer hvilke humlearter som kan finnes i et område, er tilgangen på blomster og bolplasser trolig de viktigste faktorene som avgjør hvor mange humler som kan finnes der. Humlebestandene påvirkes av en rekke ulike faktorer, men når blomstertilgangen er liten, kan konkurranse være en begrensende faktor. Det kan oppstå konkurranse mellom humler av samme art, mellom ulike humlearter eller mellom humler og andre arter som bruker de samme ressursene. Enkelte ganger kan det være flere humler og andre insekter på ett og samme blomsterhode. Da kan man se at humlene kan jage hverandre, eller at for eksempel dagsommerfugler og humler jager hverandre.
Som en tilpasning til næringsplantenes blomstringstid, er kløverhumle og slåttehumle av de siste humlene som kommer frem om våren. Denne tilpasningen er ikke nødvendigvis så smart: humlene må bygge opp større fettreserver for å kunne overleve vinteren, de får en kortere sesong og kolonier med færre individer, samt at de får tøffere konkurranse om næringsplanter fra andre insekter og humlearter som allerede har etablert bol og fått frem de første arbeiderne når kløverhumledronningen våkner. Å starte sesongen seint reduserer også mulighetene for å finne en egnet bolplass fordi mange av de beste lokalitetene allerede da er okkupert av andre humler.
Med sine lange tunger, er humlene spesialister på å hente nektar fra blomster med dype og smale kronrør. De ulike humleartene har ulik tungelengde, og dette er et trekk man antar har utviklet seg som et resultat av konkurranse mellom dem. Dersom de ulike humleartene deler på ressursene gjennom å utnytte forskjellige plantearter, kan flere humlearter sameksistere i større bestander innenfor samme areal. Arbeidere og dronninger vil ha ulike konkurranseegenskaper blant annet på grunn av forskjeller i størrelse og tungelengde. Dersom det er kamp om en attraktiv ressurs, vil de store kunne jage bort de små. Størst konkurranse vil det trolig være mellom de langtungete artene, da blomsterutvalget for slike ofte er begrenset. Man kan for eksempel tenke seg at hagehumle, som er en vanlig langtunget art, kan konkurrere direkte om ressursene med de mer sjeldne artene kløverhumle og slåttehumle.
Siden humlene trenger godt med blomster gjennom hele sesongen, vil konkurranse kunne oppstå særlig i perioder med lite blomster. Dette gjelder spesielt tidlig på våren dersom nøkkelarter som selje mangler, eller om sommeren etter at store blomsterenger er slått. Om høsten, når de fleste blomstene er avblomstret, kan det også være ei vanskelig tid.
Ved utsetting av honningbier kan man indusere en konkurransesituasjon med andre pollinatorer (Dupont et al. 2015). Honningbier unngår imidlertid normalt planter med lange kronrør, men siden de er generalister og kan opptre i store mengder, kan de også utnytte mer marginale planter som er viktige for ville bier. Honningbier går ofte på enkeltressurser i optimal blomstring, og vil da kunne fortrenge spesialister på slike planter. Trolig øker pollineringen ved utsetting av honningbier fordi de enkelte biene blir jaget og bruker kortere tid per plante og besøker derfor flere planter. I hvilken grad honningbier påvirker bestandene av humler og andre villbier er lite kjent, men de fleste studier viser at tettheten av villbier reduseres som følge av honningbier uten at dette nødvendigvis går ut over diversiteten av bier (Forup & Memmott 2005).
I kløveråkrer er det observert lavere tetthet av humler der det er satt ut honningbier (Wermuth & Dupont 2009). Det er kjent at produsenter av rødkløverfrø ofte har satt ut bikuber i håp om å bedre frøproduksjonen. Selv om honningbiene i liten grad klarer å utnytte nektaren, så kan de sanke pollen slik at det blir mindre tilgjengelig for humlene. Og når biene er mange, fjerner de også såpass mye nektar at det tar lengre tid for humlene å finne nok. Resultatet kan bli at humlene ikke klarer å produsere store nok bol, noe som igjen kan gå utover dronningproduksjonen.
I Norge på 1990-tallet begynte man å bruke bol av mørk jordhumle Bombus terrestris i veksthus for pollinering av tomater. Det har siden vist seg at enkelte nye dronninger rømmer fra veksthusene og etablererer seg i naturen. Dette har trolig vært medvirkende til den store ekspansjonen av mørk jordhumle i Norge. Oppdrettet har inntil 2013 vært basert på dronninger fanget i Norge av den hjemmehørende underarten B. t. terrestris.
Mørk jordhumle har flere konkurransefordeler sammenlignet med de fleste andre humlearter. Den er blant de aller tidligste artene om våren og vil kunne okkupere bolplasser før andre arter, den er stor og kraftig og produserer større bol med flere arbeidere og kjønnsdyr enn andre nærstående arter, og den viser stor tilpasningsdyktighet i ulike klimasoner og habitater. I tillegg er den lite kresen i matveien og er i stand til å hente næring langt fra bolet, samt at den har få naturlige fiender (Dafni et al. 2010). Når den betjener blomster med dype og smale kronrør, for eksempel rødkløver, er det ofte at den biter hull nede på kronrørene for å få tak i nektaren. Dette resulterer i mindre effektiv pollinering og dessuten at arten konkurrerer med kløverhumle og slåttehumle om deres favorittblomst, rødkløver.
Det er derfor all grunn til å tro at mørk jordhumle kan etablere seg nye steder og konkurrere om både bolplasser og blomsterressurser med stedegne humlearter også i Norge. I tillegg frykter man at importerte humler kan overføre parasitter og smittsomme sykdommer og blande seg med hjemmehørende bestander av mørk jordhumle etter at det fra 2013 ble åpnet for import av utenlandske bestander av mørk jordhumle som dels tilhører en annen underart; B. t. dalmatinus. Fra Storbritannia frykter man spesielt at importhumler kan utkonkurrere den hjemmehørende britiske underarten av mørk jordhumle B. t. audax (Ings et al. 2006).
Alt i alt er det mye som tyder på at konkurranse internt mellom humler eller mellom humler og honningbier kan være betydelig når ressursen det konkurreres om er begrenset. Slik konkurranse er imidlertid vanskelig å påvise fordi forskjeller i artssammensetning over tid også kan skyldes en rekke andre forhold. Ikke minst er det vist at når artene utsettes for flere stressfaktorer samtidig, kan effekten bli større enn summen av enkelteffektene (González-Varo et al. 2013). I et landskap der pollinatorene utsettes for tap av leveområder, giftige plantevernmidler, klimaendringer og spredning av sykdommer, vil derfor økt konkurranse være en betydelig tilleggsfaktor som del av det store bildet.

Lys jordhumle Bombus lucorum, honningbie Apis mellifera og blomsterflue i samme matfat.
Fiender og sykdommer
Humler er bytte for forholdsvis få predatorer. De fleste edderkoppspinn er for spinkle til å fange humler. Noen edderkopper som sitter gjemt i blomster kan derimot fange humler. Fugler unngår som regel humler på grunn av giftbrodden, men særlig tornskate, vepsevåk og fluesnappere kan ta humler. Det er også observert at meiser, stær og steinskvett har tatt humler. Grevling, mår, mink, rev og spissmus kan også ta humler. Grevling og bjørn er også kjent for å grave opp humlebol.
En del insektarter er kjent for å være parasitter på humler. Eksempler er parasittiske fluer som vepseflua Sicus ferrugineus, blomsterflua Volucella bombylans og spyflua Brachiocoma devia som legger sine egg i humlebol. Deres larver lever på humlenes larver og pupper. Særlig plagsom for humlene er nattsommerfuglen humlebolvoksmott,Aphomia sociella som spiser nesten alt av organisk materiale i bolet.
Det finnes også et stort antall virus, bakterier, sopp, protozoer, midd og rundormer som er tilknyttet humler. En amerikansk undersøkelse har vist at noen humlearter har gått tilbake med opptil 96 % i løpet av de siste 20 år (Cameron et al. 2011). En medvirkende årsak er trolig den parasittiske soppen Nosema bombi (Microsporidia).
Nylig er det vist at humler rammes av sykdomsvirus som tradisjonelt er knyttet til honningbier og at dette er et nokså vanlig fenomen. Det samme gjelder også for smitte fra oppdrettshumler til stedegne humler (Meeus et al. 2011). Først ble det kjent at vingedeformasjonsvirus (DWV) kan overføres fra honningbier til humler (Fürst et al. 2014), og nylig er det også dokumentert at fire andre sykdomsvirus tilknyttet honningbier kan smitte til humler (McMahon et al. 2015). Hvordan dette smitter er lite kjent, men en teori er at det det skjer via blomsterbesøk og pollen.

Til tross for varselfarger og stikkebrodd har humlene mange fiender. Her har en krabbeedderkopp tatt ei åkerhumle. Flere arter av pattedyr og fugler tar også humler, men av størst betydning for regulering av humlesamfunnene er trolig parasitter og sykdomsorganismer, samt klima og værforhold.
Genetikk
Humler har som andre sosiale Hymenoptera-arter et haplo-diploid genetisk system, hvor hunnene (dronninger og arbeidere) er diploide (dobbelt sett av kromosomer), mens hannene er haploide (enkelt sett av kromosomer).
Dersom humlebestander reduseres til et minimum, vil de bli mer sårbare for tilfeldige hendelser. Humlesamfunnet er grunnlagt av en enkelt dronning, og på grunn av det haplo-diploide genetiske systemet blir den effektive bestandsstørrelsen lik antall humlebol multiplisert med 1,5. Små isolerte bestander vil ha liten eller ingen utskifting av individer med nabobestander, og vil dermed tape genetisk mangfold. Dette kan føre til at skadelige recessive (vikende) alleler (alternative utgaver av et gen) får innpass i bestanden. Små bestander kan være utsatt for innavlsdepresjon. Det er påvist redusert kolonidannelse, lavere eggproduksjon hos dronninger og redusert vinteroverlevelse som resultat av paring mellom bror og søster.
Et annet eksempel på innavlsdepresjon er at det i små bestander kan opptre reduksjon i antall kjønnsalleler, noe som øker sannsynligheten for at en dronning parer seg med en hann som har en av hennes kjønnsbestemmende alleler. Det kan resultere i at det produseres 50 % diploide hanner. Disse hannene har lav fertilitet og redusert immunrespons, og representerer derfor en kostnad for kolonien. Diploide hanner er sjelden registrert under naturlige forhold, men er påvist hos artene kysthumle, enghumle og lynghumle i Skottland (Ellis et al. 2006, Darvill et al. 2012). Det ble i samme området påvist triploide arbeidere av kysthumle og lynghumle, noe som viser at diploide hanner er fertile (Darvill et al. 2012).
Innavl fører ikke nødvendigvis til innavlsdepresjon og at bestander dør ut. Det er mange eksempler på at introduserte humlebestander har klart seg godt selv om de er grunnlagt av få utsatte individer. Selv om det ikke foreligger sikre bevis på at innavl har ført til at små isolerte bestander av humler har dødd ut, er det likevel sannsynlig at det ofte har vært en medvirkende årsak (Goulson et al. 2008). I en undersøkelse i Nord-Amerika ble det funnet at humlebestander med stor bestandsnedgang (opptil 96 %) hadde lav genetisk diversitet og høyere infeksjonsnivå av sykdomsparasitten Nosema bombi sammenlignet med arter som hadde stabile bestander. Det ble antatt at økt innavl og genetisk drift i små bestander ga økt mottagelighet for infeksjon av denne sykdomsparasitten (Cameron et al. 2011).
For å sikre langsiktig bevaring av våre mest sjeldne humler er det viktig at leveområdene er store nok og at delbestander har forbindelse med hverandre slik at de kan utveksle individer og derved redusere risikoen for innavl.
Endringer i kulturlandskapet
Det har skjedd store endringer i kulturlandskapet i Norge de siste 150 år, noe som utvilsomt har hatt betydning for humlebestandene. På den ene siden har landbruksdriften blitt intensivert i noen områder, mens på den andre siden er gjengroing en pågående prosess i store deler av landet. Et resultat av begge disse prosessene er at blomstertilbudet for humlene har blitt redusert. I det intensivt drevne jordbrukslandskapet har både mengde og mangfold av de planteartene som mange humlearter foretrekker blitt redusert. De gamle slåtteengene, samt de blomsterrike og ugjødslete beitemarkene, er mange steder nedbygd eller erstattet med monokulturer. Bruken av kunstgjødsel har også medført at kulturlandskapet er mindre attraktivt for humlene med dominans av gras og konkurransesterke planter. Selje og vierarter (Salix spp.) er viktige næringsplanter for humlene tidlig om våren når samfunnene grunnlegges. Imidlertid er humlene avhengige av et kontinuerlig tilbud av ulike plantearter som pollen- og nektarkilde utover hele sommeren og høsten. Dersom deler av sesongen har lite blomster, vil det kunne påvirke humlebestandene gjennom hele året.
Mangel på egnete bolplasser kan være en viktig årsak til reduserte humlebestander. De fleste av våre humlearter plasserer bolet oppå eller nede i bakken, gjerne i et gammelt smågnagerbol eller i moselaget. I jordbrukslandskapet finnes humlebolene langs åkerkanter, i gressbakker, langs vann og vassdrag og på åkerholmer. Humlene krever åpenmark som i liten grad er nylig bearbeidet, noe som kan være vanskelig å ta hensyn til med dagens jordbrukspraksis. Arealer med mye smågnagere er ekstra gunstige, og humler kan til og med fordrive smågnagere fra bolene sine. Brenning av åker, eng, skråninger og kantsoner kan også være årsak til tap av bolplasser gjennom at musebol og løst gress som humlene kunne brukt blir brent opp, samt at beskyttende vegetasjon blir borte.
Bruk av ulike sprøytemidler (pesticider) i hager, åkre og kantsoner kan påvirke humler. Det er dokumentert at enkelte stoffer innenfor sprøytemiddelgruppen neonikotinoider i realistiske doser, kan redusere kolonitetthet, kolonivekst og reproduksjon hos villbier inkludert humler i naturen (Goulson 2015, Rundlöf et al. 2015). Stoffene påvirker blant annet bienes orienteringsevne og dronningproduksjonen hos humler (Whitehorn et al. 2012). Effektene av pesticider kan både være direkte, gjennom at humlene forgiftes, eller indirekte ved at blomsterplanter forsvinner. Sprøytemidler har trolig størst negativ effekt om våren da bolene er små og dronningene er ute for å samle næring. Når koloniene er store, kan de bedre tolerere tap av noen arbeidere.

Blomsterrike slåtteenger er viktige naturtyper for humler og en rekke andre insekter. I dag er det langt mellom slike områder, og mye tyder på at mangel på blomster er en viktig årsak til at flere arter av pollinerende insekter har gått tilbake. Bildet er fra Aremark i Østfold.
Klimaendringer
Klima er en av de viktigste faktorene som bestemmer artenes utbredelse, enten direkte eller indirekte. Endringer i klima kan påvirke artens forekomst enten gjennom økt frekvens og styrke på ekstreme klimahendelser eller gjennom gradvise endringer av gjennomsnittlige forhold. Det finnes forholdsvis lite kunnskap om hvordan klima påvirker humler, men det er stor enighet om at klimaendringer er en viktig årsak til at enkelte humlearter er på tilbakegang og at andre er på framgang. I tillegg vil værmessige forhold forklare mye av svingningene i humlebestander fra år til år.
Tilbakegang i humlebestander forklares ofte med endringer i landbrukspraksis som reduserer ressurstilgangen for humlene. Omfattende arealendringer, bruk av plantevernmidler og kunstgjødsel vil i mange tilfeller resultere i færre bolplasser og færre blomster for humlene. Det er imidlertid lite kunnskap om hvorfor noen arter tilsynelatende blir mer sårbare enn andre når ressurstilgangen reduseres. Lenge trodde man at spesialisering i valg av vertsplanter var den viktigste årsaken til at humlearter gikk tilbake, men denne hypotesen står nå svakt alene (Williams 2005). Det kan derimot se ut til at en kombinasjon av plantepreferanser og klima kan være en viktig forklaring. Arter som har mindre toleranse for variasjon i klima ser ut til å være mest sårbare, og bestander som lever i ytterkanten av sitt klimatiske toleranseområde ser ut til å forsvinne først. Omfanget av klimaeffektene påvirkes sterkt av ressurstilgangen for humlene slik at disse effektene slår inn tidligere når blomstertilgangen reduseres. Det vil si at dersom det er rikelig med blomsterenger og leveområder for humlene, vil humlene være mer robuste for klimaendringer. Denne sammenhengen er vist for kløverhumle B. distinguendus og enghumle B. sylvarum i England (Williams et al. 2007). Klimaendringene vil med andre ord kunne slå ut bestander som allerede er svekket pga. andre påvirkninger.
Gjennom et stort EU-prosjekt har man nå samlet mer enn én million humleobservasjoner i Europa for å beregne klimatoleransen for de 69 humleartene i denne verdensdelen. Basert på tre ulike scenarier for klimaendringer fram mot år 2050 og år 2100, har man modellert hvordan disse artene vil være utbredt i framtida (Rasmont et al. 2015). Avhengig av hvilke klimascenarier og spredningsmodeller man legger til grunn, vil mellom 9 og 34 humlearter være ekstremt utsatt og står i fare for å forsvinne helt fra Europa innen 2100. I tillegg viser beregningene at mer enn 41 % av artene i Europa vil innskrenke sitt nåværende utbredelsesområde med mellom 50 og 70 % i samme periode. Kun tre arter vil kunne bli mer vanlige som følge av klimaendringene. I hvilken grad artene vil overleve i framtida, vil dels avhenge av hvordan de er i stand til å spre seg til nye områder som har mer optimale klimatiske forhold. I Norge vil det for eksempel kunne bli passende klima for arter som ikke lenger vil klare varmere og tørrere somre lenger sør i Europa. Artenes spredningsevne vil da være avgjørende for deres framtidige overlevelse. Man har allerede sett at tilpasningsdyktige arter som steinhumle B. lapidarius og mørk jordhumle B. terrestris nå har spredt seg til nord for polarsirkelen (Martinet et al. 2015), mens andre arter trekker seg tilbake.
Klimaendringene vil ganske sikkert medføre store endringer i utbredelsesmønster for humler også i Norge. Reduksjon i antall soltimer og økt nedbør om sommeren under larveutviklingen og den mest hektiske flygetiden kan være negativt, men dette vil til en viss grad kunne kompenseres av økt gjennomsnittstemperatur. En temperaturøkning uten økning i skydekke vil derimot kunne virke positivt på utviklingen av flere humlebestander, og man kan forvente at artene ekspanderer om det finnes tilstrekkelig habitattilgang. Vi ser allerede nå at fjellhumlene (underslekten Alpinobombus) opptrer mer enn 100 høydemeter lenger opp i fjellet enn bare for noen tiår siden. Dersom disse endringen skjer fortere enn vegetasjonen klarer å følge etter, kan det bli problematisk med hensyn til ressurstilgangen. I lavere fjellområder vil artene forsvinne når de har nådd toppen av fjellet og det ikke lenger er noe sted å flykte til.
Forandringer i klima vil også kunne medføre endringer som har indirekte innvirkning på humlenes livsvilkår. Blant annet forandringer i flora, artssammensetning, konkurransesituasjon og forekomsten av parasitter og sykdommer. Klimaendringer kan også være medvirkende for at frekvensen og omfanget av forstyrrelser som skogbrann og stormfellinger, som kan ha betydning for humlenes livsvilkår, ender seg. Musebol er meget viktig for de fleste arter av humler. Bestanden av smågnagere er gjerne sterkt påvirket av klimatiske forhold, og slikt som barfrost og manglende subnival (luftrom mellom bakke og snø) vil begrense bestandene betydelig. Også for humledronninger i dvale er de sistnevnte værmessige/klimatiske forhold ugunstige.
Det er vist at endringer i klima kan føre til dårligere synkronisering mellom planter og insekters aktivitetsperioder. Slike såkalte «fenologiske mismatch» kan oppstå dersom en humle, som følge av endringer i klima, forskyver sin aktivitetsperiode i tidsmessig forhold til sine næringsplanter som kan være mindre påvirket av klima (Totland et al. 2013). Dette er først og fremst et problem som rammer mer spesialiserte arter enn humler, men i perioder når få næringsplanter er tilgjengelige, som på våren, kan dette være kritisk også for humler. En annen tilsvarende vær- og klimarelatert situasjon er hvis sesongen starter ekstremt tidlig og både humler og blomster (selje) lokkes ut i begynnelsen av mars pga. uvanlig varmt vær. Humledronningene kommer da godt i gang, men står i fare for å bli utsatt for lange og kalde perioder før våren setter inn for fullt. De risikerer da å gå tomme for energi og fryse ihjel dersom bolet allerede er startet opp.

Det er stor enighet om at klimaendringer er en viktig årsak til at enkelte humlearter er på tilbakegang. Bildet viser nedsmelting av Bøyabreen i Sogn og Fjordane.
Fremmede planter
Fremveksten av fremmede planter kan ha ulike effekter på humlebestander. Faktisk er det slik at flere av de mest invaderende fremmede planteartene er populære for humler. Fremmede planter som valurt Symphytum spp., ormehode Echium vulgare, lupin Lupinus spp., steinkløver Melilotus spp., kanadagullris Solidago canadensis, russekål Bunias orientalis og ikke minst kjempespringfrø Impatiens glandulifera kan være gode trekkplanter for humler. Det er heller ikke utenkelig at plantenes fremmarsj kan sees i sammenheng med vellykket pollinering og god frøsetting. I tillegg kan noen av disse plantene også bidra til at det blir flere humler der blomsterressurser er minimumsfaktor for bestandsstørrelsene til humlene. En art som hagehumle, som har gått noe tilbake de siste tiårene, ser ut til å ha fått et oppsving igjen i områder med mye kjempespringfrø, som med sine dype blomster er en svært populær plante for denne langtungete humla.
Til tross for at fremmede planter kan være gode trekkplanter for humler, er mange av dem sterkt uønsket i naturen. Trolig har de også flere negative effekter på humlesamfunnene. For det første fortrenger disse plantene andre gode trekkplanter for humlene slik at arter som er mer avhengige av hjemmehørende planter vil få reduserte muligheter. Flere av de fremmede plantene gir mat kun en kort periode i sesongen. Det er heller ikke kjent i hvilken grad arealer med fremmede planter påvirker potensialet for bolplasser til humler.
Den fremmede arten lupin er mange steder en viktig pollenleverandør for flere humlearter, også for fåtallige arter som kløverhumle og slåttehumle. Lupinblomstene leverer imidlertid ikke særlig mye nektar. Det er også vist at pollen fra lupiner kan inneholde antibeitestoffer som kan påvirke humlenes reproduksjonsevne (Arnhold et al. 2014). Det er imidlertid ukjent i hvilken grad slike påvirkninger er et mer generelt trekk med fremmede planter og i hvilken grad dette gjelder under lokale forhold der de fremmede plantene er lite belastet med herbivorer som induserer produksjon av antibeitestoffer.
Selv om mange hageplanter er sterkt uønsket i naturen, er det ingen tvil om at blomsterrike hager kan være viktige habitater for mange humler. Med økende bevissthet rundt dette med å ta vare på pollinatorer, har mange begynt å plante humlevennlige planter i parker og hager. Dette kan være et viktig tiltak lokalt og spesielt vår og høst da det kan være lite blomster å finne i naturen. En særdeles nyttig plante for mange langtungete humler er sibirertebusk Caragana arborescens. Denne blomstrer om våren når humledronningene trenger mye næring og kan trekke enorme mengder humler fra store områder. En plante som blomstrer seint, og som er viktig for langtungete humler, er blomkarse. Den kan være viktig for lushatthumle etter at tyrihjelmen er avblomstret.

Dronning av kløverhumle Bombus distinguendus. Lupiner kan være en god pollenkilde for humler, men planten er svartelistet og uønsket fordi den fortrenger stedegne plantearter og endrer naturtypene der den vokser. Dette kan også være negativt for humlesamfunnene.
Giftige planter og biltrafikk
Humlebolene er ettårige, og det er kun de befruktete dronningene som overvintrer. Store deler av årets humleproduksjon vil derfor dø i løpet av sesongen, men selv om dette er nokså store insekter, er det relativt sjelden å se døde humler i naturen. Av og til kan man imidlertid finne store mengder døde humler under visse arter av blomstrende trær og langs veikanter. Humledød under blomstrende lind har særlig vært mye diskutert, men humledøden langs veiene våre er trolig vel så omfattende. Det er imidlertid usikkert hvilken betydning slike trusselfaktorer har for bestandene, og hvilken betydning det har sammenlignet med de mer langstiktige arealendringene.
Det observeres jevnlig store mengder døde humler under enkelte blomstrende lindetrær. En av de tidligste teoriene for å forklare dette fenomenet gikk på at nektaren inneholder sukkertypen mannose, som man trodde var dødelig for humler og bier selv i små konsentrasjoner. Senere har det vist seg at dette trolig ikke stemmer. En annen teori går ut på at humlene dør av sult ved at blomstene er gått tomme for nektar og pollen, men at blomstene fortsetter å skiller ut sterke dufter som humlene ikke klarer å motstå. Andre teorier går på at det kan ha sammenheng med gjæring og uttørking. Observasjoner av slik massedød er gjort særlig i tilknytning til fremmede og hybridiserte lindearter og lønn, men også hos fremmede bartrær.
Kollisjoner med kjøretøy langs sterkt trafikkerte veier er en annen trussel for humlebestandene. Gjennom hele humlesesongen finner man store mengder døde humler i veikantene. Langs veiene er det ofte et rikt mangfold av planter som trekker humler, og på mange måter kan vi si at de tidligere blomsterrike slåtteengene er flyttet til veikantene. Enkelte steder kan derfor en stor andel av humlebestandene i et område søke nektar på blomster i veikanter. Om våren, når seljene står i blomst, finnes store mengder av humledronninger på disse. Selje etablerer seg raskt i veikantene, og trær som blir felt produserer stubbeskudd som i løpet av få år gir store mengder seljeblomster.
Selv om store mengder humler dør under lindetrærne og i biltrafikken, har dette kanskje mindre betydning for bestandene enn man skulle tro? Lind blomstrer på et tidspunkt da humlebestandene er på topp og de fleste arbeiderne likevel snart skal dø, og humledøden langs veiene er kanskje ikke større enn at bestandene opprettholdes på noenlunde samme nivå som uten trafikk? Men dersom store mengder humlearbeidere dør et par uker tidligere enn normalt, kan dette ha vesentlig betydning. På denne tiden er det kjønnsindividene i humlebolet som fostres opp, og et frafall i arbeiderstokken kan derfor ha stor indirekte betydning på produksjonen av nye dronninger og hanner. De nye dronningene, som klekkes på ettersommeren, er ute av bolet en kort periode for å pare seg og ta til seg opplagsnæring til overvintringen. Dersom disse dronningene tiltrekkes av lind og lider samme skjebne som arbeiderne, vil det være særlig alvorlig for bestanden. Hvis humledronninger dør i trafikken om våren, vil som regel hele bolet gå under, men etter at arbeidere er produsert, kan en arbeider delvis ta over dronningfunksjonen hvis dronninga faller fra. Disse kan imidlertid bare produsere hanner ut av egne egg.

Veikanter har ofte et rikt mangfold av planter som utgjør et viktig tilskudd av pollen og nektar for humlene. Ulempen er imidlertid at humlene risikerer å bli påkjørt i trafikken. Bildet viser setermjelt langs fylkesvei 29 gjennom Folldal i Hedmark.
Humler som modeller
Mange humler ligner hverandre sterkt i fargetegningene. Dette er en av årsakene til at det ofte er vanskelig og usikkert å artsbestemme humlene på basis av slike mønstre, men det er faktisk en fordel for humlene å ligne på hverandre. Når flere giftige arter (eller arter med giftbrodd som humler) ligner på hverandre, kan hvert enkelt individ dra fordel av det ved at man lettere unngår å bli spist av insektetere. Dette fenomenet kalles «Müllersk mimikry» eller «mimikrykomplekser». Det er også vanlig at humler kan ha et spekter av ulike fargemorfer (en variant av en art med et bestemt fargemønster) med større variasjon i fargemønster innen en art enn mellom arter. Av humler i Norge er enghumle det beste eksempelet på dette. Den arten opptrer i en mørk form og i en lys form. For å gjøre det ekstra vanskelig for oss som prøver å artsbestemme humlene, er det slik at ulike arter kan ha et identisk variasjonsspekter i fargemorfer. Hagehumle kan i en melanistisk form ligne helt på våre slåttehumler. Samtidig opptrer ofte individer av ulike arter med identisk fargemorf på samme sted. Dette ser vi blant annet hos berghumle og lapphumle.
Siden humler kan være giftige for insektetere, vil spiselige dyr som ligner humler kunne ha en fordel ved at de sjelden blir spist. Dette er bakgrunnen for at en rekke andre insekter ligner sterkt på humler selv om de er fjernt beslektet. Fenomenet kalles «batesisk mimikry» og er svært vanlig i naturen. Hos enkelte insektgrupper er slik mimikry mer utpreget enn hos andre. Det er særlig hos insekter som lever i de samme miljøene og som har en genetisk disposisjon til det, at mimikry er spesielt vanlig.
I Norge står blomsterfluene (Syrphidae) i en særstilling og inkluderer en rekke arter som etterligner både humler, andre bier og stikkeveps. Den vanligste og kanskje mest spesielle av disse er humleblomsterflua Volucella bombylans. Denne kan opptre i flere ulike fargemorfer som minner om hver sine grupper av humler. Det finnes både jordhumle-, steinhumle-, og trehumlelignende varianter. Ikke nok med at den etterligner humlenes farger og atferd gjennom summing og bevegelser, men når flua sitter på en blomst kan den strekke frambeina framover langs hodet slik at det ser ut som den har kraftige humleantenner (se bilde). Arten utvikler seg i humlebol der den lever av rester og avfall, men den kan også ta humlelarver. Den har derfor ikke bare behov for å beskytte seg mot fiender, men også for å kamuflere seg mot humlene selv.
Totalt sju av våre blomsterfluearter er typiske humleetterlignere. Bjørneblomsterflua Arctophila bombiformis har jordhumlemønster og er kanskje den største av dem alle. Den er ikke uvanlig i skogkanter på Østlandet og i Trøndelag om sommeren. Loddenblomsterflue Pocota personata er en mer sjelden og litt mindre art med jordhumlemønster som er knyttet til løvskogsområder på Østlandet. Svart pelsblomsterflue Criorhina ranunculi er en bemerkelsesverdig art som ofte er å finne sittende på trestammer, gjerne osp, tidlig om våren. Denne vakre blomsterflua etterligner de svarte humlene med rød bakstuss og finnes spredt nord til Trøndelag. En annen art i sammen slekt, Criorhina berberina, er sjelden og kun påvist et par ganger i Norge. Denne opptrer i to fargeformer; en som ligner jordhumler og en som ligner trehumle. Ellers finnes ytterligere et par humleetterlignere med lyse tegninger på brystet og rød bakstuss: praktdroneflue Eristalis oestracea og humlebarskogflue Erizona syrphoides som begge er relativt vanlige hos oss.
Det er også en del andre tovinger (Diptera) som kan minne om humler. For eksempel den store rovflua Laphria flava og møkkflua Mesembrina mystacea. Sistnevnte er en utpreget humleetterligner, og minner mest om trehumle eller barskoghumle. Det passer fint for en art som er mest vanlig i skogen. Blant snyltefluer (Tachinidae) finnes også noen store og kraftige arter som kan minne om humler, for eksempel den helsvarte Tachina grossa og den mer oransjetegnete Tachina fera, selv om disse ikke har typiske fargemønster som humler. Innenfor den parasittiske fluefamilien brems (Oestridae) finnes også flere arter som ligner humler. I fjellet kan både reinbrems Hypoderma tarandi og reinens nesebrems Cephenemyia trompe minne om (særlig hanner av) berghumle og lapphumle.
Når det gjelder andre arter blant årevinger (Hymemoptera), er det i grunnen ikke så mange humleetterlignere, selv om de store klubbevepsene (Cimbicidae) i slektene Trichiosoma og Cimbex, både i størrelse og farger, kan ha likhetstrekk med humler. Også andre villbier kan ligne humler. Blant disse er humlepelsbie Anthophora furcata en åpenbar kandidat til å bli feilbestemt som for eksempel en enghumlearbeider. Noen gjøkhumler ligner sine verter, slik som steingjøkhumle og jordgjøkhumle, men dette er ikke et typisk trekk for andre gjøkhumler som markgjøkhumle og tregjøkhumle. Kanskje er ikke pelsfarge så viktig for å unngå å bli avslørt som gjøk i humleverdenen? Det er likevel spekulert i om årsaken til at lundgjøkhumle har rød bakstuss, i motsetning til verten lundhumle, skyldes at lundgjøkhumle opprinnelig er tilpasset underarten proteus av lundhumle som har rød bakstuss og finnes lenger sør.
Også blant sommerfuglene finner vi humleetterlignere i flere familier. Glassvingene (Sesiidae) er trolig de mest utpregete der alle arter ligner mer på andre insekter enn sommerfugler. For eksempel ospeglassvinge Sesia apiformis, som med sin tykke bakkropp og gule og svarte tegninger noen ganger blir forvekslet med ei humle. Det er imidlertid en stor stikkeveps, muligens geithams Vespa crabro, som har vært modell for denne. Gulkrageglassvinge Sesia bembiciformis, er en annen, meget sjelden art på samme størrelse og med lignende fargetegninger.
Ellers er humlesvermerne kanskje de aller beste etterlignerne blant sommerfuglene. De tilhører familien tussmørkesvermerne (Sphecidae), men er dagflygende. Vi har to arter i Norge: smalkantet humlesvermer Hemaris tityrus, som finnes nord til Trøndelag, og bredkantet humlesvermer Hemaris fuciformis, som er mer begrenset utbredt på Sørøstlandet.
Man kommer ikke utenom å nevne humlebille Trichius fasciatus. Selv om det er lett å se at dette er ei bille, har den humlelik behåring og fargetegninger som gjør at det er mulig å ta feil ved første øyekast. Særlig når den flyr og nærmer seg en blomst med sin summende og humlelike atferd skal det mer til å avsløre forkledningen.

Humleblomsterflua Volucella bombylans er en av de mest vellykkete etterlignerne av humler. Her strekker den frambeina langs hodet for tilsynelatende å få antenner som ei humle.

Gulstripet bjørneblomsterflue Arctophila bombiformis har fargemønster som ei jordhumle.

Humlepelsbie Anthophora furcata er en av våre største villbier og er svært lik en humle.

Reinens nesebrems Cephenemyia trompe samles ofte på fjelltoppene og kan minne om ei lapphumle. Finnshøa i Oppdal, Sør-Trøndelag.

Humlebiller Trichius fasciatus på hvitbladtistel, Munkvoll, Oppdal i Sør-Trøndelag.

Den store rovflua Laphria flava kan også ligne på en humle.
Kartlegging av humler i Norge
Kunnskapen om norske humler er unik gjennom arbeidene til Astrid Løken (1911-2008) gjennom flere tiår (Løken 1973, 1984). Vi har derfor en solid basis for å vurdere endringer i humlefaunaen i Norge. De siste årene har vi sett en enorm økning i interesse for humler, slik at oppdatert kunnskap om både utbredelse og bestandsstatus for humler i Norge må sies å være relativt god. Humlene ble for første gang rødlistevurdert i Norsk Rødliste for arter 2010, der arbeidet med humler i stor grad var basert på statusoversikten i Ødegaard et al. (2009). Et nytt og mye større kunnskapsgrunnlag ble lagt til grunn for de reviderte rødlistevurderingene av humler for Norsk Rødliste for arter 2015 (se referanseliste for nyere kartleggingsrapporter).
I forbindelse med den nasjonale satsingen Naturindeks for Norge ble det startet omfattende humlekartlegging i seks fylker i Norge. Østfold og Vestfold startet i 2009 (Öberg et al. 2010), og er siden utvidet til Sør-Trøndelag, Nord-Trøndelag, Vest-Agder og Rogaland (Öberg et al. 2011, 2012, Åström et al. 2013, 2014). Humler har ellers vært en vesentlig del av store kartleggingssatsinger gjennom Artsprosjektet (prosjektene INVENT-ART 2010–2013 og ACUNOR 2014–2016). Kløverhumle, slåttehumle, bakkehumle og lundgjøkhumle har dessuten vært gjenstand for særlig kartleggingsfokus gjennom deres status som rødlistearter og handlingsplanarter i regi av Fylkesmannen i Oslo og Akershus (Ødegaard et al. 2013, Gjershaug et al. 2013, Bengtson & Olsen 2013a,b, Bengtson & Olsen 2014).
Ellers er det verdt å merke seg at humlene har fått enorm oppmerksomhet i media de siste årene. Dette har skapt økt fokus på humler både blant allmennheten, samt i forskning og forvaltning. Denne økte interessen har medført økt kartleggingsaktivitet på alle plan. Mens det i perioden 1975–2005 knapt ble registret ei humle i Norge, representerer det tilsvarende antallet registrerte observasjoner i neste tiårsperiode mer enn 13 000 poster i Artsdatabankens tjeneste Artskart.

Interessen for kartlegging av humler har økt enormt de siste årene. På bildet registreres arter og antall individer i faste flater som en del av overvåkingen av humler i Norge. Slike data kan brukes til å beregne hvordan det går med bestander av vanlige humlearter over tid.
Bestemmelse av humler
Bestemmelse av humler kan være både svært enkelt og veldig vanskelig, alt etter hvilke arter/kaster det er snakk om og hva slags materiale man har tilgjengelig. Mange arter kan opptre i form av atypiske individer, og det er ofte vanskeligere å bestemme arbeidere enn dronninger. Hanner byr på særlige utfordringer hos noen arter. Det er også betydelig vanskeligere å artsbestemme med utgangspunkt i bilder enn innsamlede individer. Noen individer er ikke mulig å artsbestemme verken basert på ytre eller indre morfologiske karakterer og bør DNA-strekkodes for sikker artsidentifisering.
En enkel start for den som vil studere humler er å få oversikt over grupper av humler basert på fargemønstre. Spesielt i lavlandet i Sør-Norge vil de fleste humlene man ser tilhøre en av følgende tre grovt inndelte fargegrupper: «jordhumle-gruppen» med tverrstriper i sort, gult og hvitt, «åkerhumle-gruppen» med brunoransje mellomkropp og «steinhumle-gruppen» med sort over det meste bortsett fra rødaktig bakerst. En rekke ulike diagnostiske karakterer kan sjekkes for en humle innenfor hver av disse gruppene, slik at en sikker artsbestemmelse kan oppnås. Fargeavvik og variasjon er ikke uvanlig hos humler, og bestemmelsesnøkler kun basert på farger vil ofte ikke gi en sikker artsbestemmelse.
Når man skal artsbestemme en humle, er det nyttig å først avgjøre hvilket kjønn den tilhører og om det er en sosial humle eller en gjøkhumle. Ofte er dette mulig å avgjøre når vi ser en humle sitte i en blomst. Tidlig om våren er det bare dronninger som er ute, men etter noen uker kommer også arbeiderne. Dronninger og arbeidere (som vi med en samlebetegnelse kan kalle "hunner") av sosiale humler samler pollen i pollenkurver på bakbeina. Bakre leggbein hos disse er brede, flate og blanke på leggflaten, men kantene av leggen har stive lange hår som danner en pollenkurv (corbicula). Dette kan være vanskelig å se i felten dersom humlene ikke har pollenklumper i pollenkurven. Gjøkhumledronningene kan vi kjenne igjen på at de har mer glissen behåring på bakkroppen slik at de ser mer glinsende ut, samt at bakre leggbein mangler pollenkurver.
På ettersommeren dukker hannhumlene opp. Disse ligner på hunnene, men er smalere i kroppsformen og har mer avrundet bakstuss. De har lengre antenner med 13 ledd, mens dronninger og arbeidere har 12 antenneledd. Hannenes fargetegninger ligner ofte hunnenes, men kan være mer variable. Hanner hos mange arter har lys behåring i ansiktet.

Artsbestemmelse av humler under stereolupe.
Kroppsbygning

Humlekroppen, overside
Humlekroppen sett ovenfra med angivelse av kroppsdeler og betegnelser som har betydning for artsbestemmelse. Bildet viser en dronning av enghumle Bombus sylvarum.

Humlekroppen, underside
Humlekroppen sett fra undersiden med angivelse av kroppsdeler og betegnelser som har betydning for artsbestemmelse. Bildet viser en dronning av enghumle Bombus sylvarum.

Humlebein
Bakbein av en hunn av en sosial humleart med navn på de ulike leddene, samt begreper som er brukt i humlelitteratur.

Humlehode, forfra
Humlehode sett forfra med angivelse av betegnelser som har betydning for artsbestemmelse. Bildet viser en hunn av steingjøkhumle Bombus rupestris.

Humlehode, fra siden
Humlehode sett fra siden med angivelse av betegnelser som har betydning for artsbestemmelse. Bildet viser en dronning av slåttehumle Bombus subterraneus.
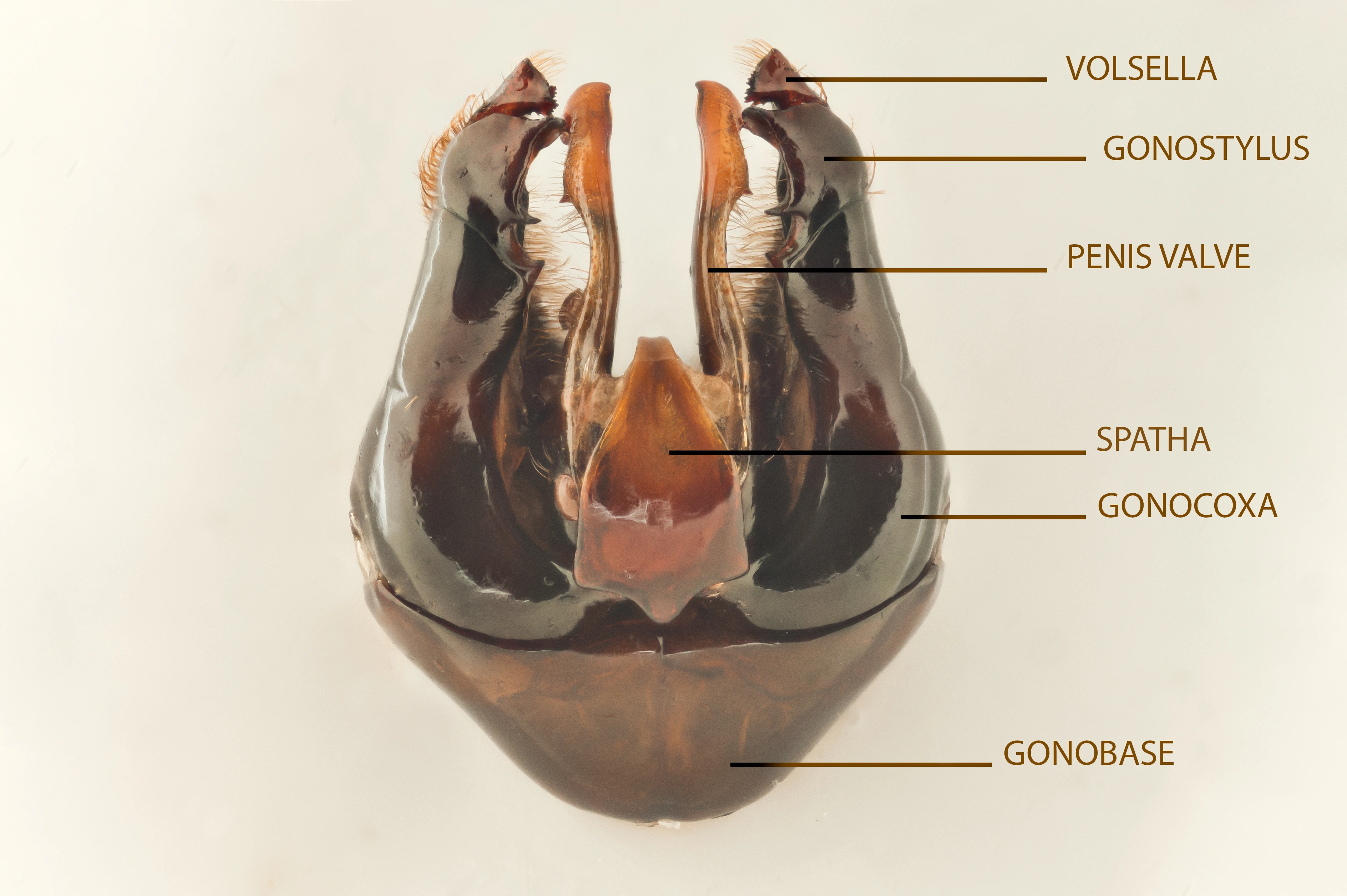
Genitalia
Hannlige genitalia av ei humle med betegnelser på de ulike delene. Bildet viser fjellhumle Bombus balteatus.
Navn på humlenes kroppsdeler og betegnelser som vil omtales i beskrivelsen av de enkelte artene og som har betydning for artsbestemmelsen er presentert her. Hodet har en rekke viktige karakterer som er avgjørende for artsbestemmelse. Spesielt er kinnlengden (eng.: malar space) avgjørende. Denne måles som forholdet mellom kinnets lengde og kinnets bredde ved basis av overkjevene (mandiblene) (se målelinjer på bildet). Munnskjoldet (clypeus) og overleppen (labrum) har også strukturer og punktering som kan være viktige. Likeledes er mandiblenes form og tungas (glossa) lengde varierende fra art til art. Hos hannene er behåringenes farge på panne (vertex) og ansikt (frons) av betydning hos flere arter. Pannens grad av punktering og blanke flater mellom fasettøye og punktøye (ocellus) kan også ha betydning for artsbestemmelsen.
Mellomkroppen (mesosoma eller thorax) er delen mellom hodet og bakkroppen der vinger og bein er festet. Det er særlig behåringens farge og mønster på mellomkroppens overside som er viktig for artsbestemmelse. Kragen eller kragebåndet er mellomkroppens fremste del som ofte har avvikende farge fra den midtre delen. Båndet mellom vingefestene kan kalles interalarbånd (ryggbånd). Skutellen er den bakerste delen av mellomkroppen, og kan ha behåring med avvikende farge som danner et skutellbånd.
Beina hos humler består av 9 ledd som ender i to klør. Fra beinfestet er leddene hofte (coxa), hoftering (trochant), lår (femur), legg (tibia), basitarsus (av og til kalt "metatarsus", men dette begrepet er noe forvirrende siden hele baktarsen også noen ganger kalles "metatarsus"), 3 tarseledd (tarsomerer) og kloledd (pretarsus). Basitarsus, tarsomerer og pretarsus utgjør de fem tarseleddene. Dronninger og arbeidere av sosiale humler har en såkalt pollenkurv på bakleggene. Hårene som danner denne kurven, har noen ganger forskjellig farge hos ulike arter. Utformingen av midtre basitarsus og bakre legg (tibia) kan være karakteristisk for enkelte underslekter, og kan dermed være viktig for å skille arter med lignende fargemønster. Ribbenettet på vingene kan også ha små forskjeller, men dette er ikke vektlagt her.
Bakkroppen (metasoma eller abdomen) består av ryggplater (tergitter) på oversiden, T1–T6 hos hunnene og T1–T7 hos hannene, og bukplater (sternitter) på undersiden (St1–St6). De tre siste ryggplatene hos hunnene (T4–T6) og de fire siste hos hannene (T4–T7) kaller vi "bakstussen" (eng.: tail). Denne har ofte annerledes farge på behåringen. Hannene har i tillegg to indre bukplater (St7 og St8) som ofte er viktige for artsbestemmelse. Hannene har komplekst bygde genitalier som gir en sikker artsbestemmelse for de fleste arter (se figur under). Hos dronninger og arbeidere er de indre bakkroppsdelene omdannet til en broddslire (eng.: sting-sheath) og en stikkebrodd. Når man samler inn hanner for montering og artsbestemmelse, er det lurt å preparere ut genitaliene, samt St7 og St8, og lime disse delene opp på en egen plate som nåles sammen med humla. På samme måte kan det være lurt å ta ut broddslira på dronninger og arbeidere hos arter der denne kan være avgjørende for artsidentifikasjon.
Praktiske tips
Humler finnes nesten overalt der det finnes blomster, og kan oftest studeres, fotograferes og artsbestemmes mens de sitter på en blomst. De fleste eksemplarer lar seg lett artsbestemme med litt trening. Noen eksemplarer er imidlertid så vanskelige eller atypiske, at de selv på gode fotos kan være vanskelige å artsbestemme. Slike eksemplarer kan det være nødvendig å samle inn.
Humler kan fanges med en insekthåv og forsiktig overføres til et lite glass for nærmere studier. De bør ikke oppbevares der i mer enn 15 minutter, helst kortere. En bør unngå å utsette glasset for sollys. Dersom individet krever undersøkelse av små detaljer med lupe, kan man bedøve humlen med CO2-gass.
Enkelte vanskelige eksemplarer må studeres under stereolupe for sikker artsbestemmelse. Det gjelder for eksempel berghumle og lapphumle. En bør ikke ta livet av dronninger om våren, særlig ikke av de fåtallige artene. Dersom en ikke klarer å artsbestemme en humledronning i felt om våren, kan en komme tilbake senere på sommeren og samle inn en arbeider eller hanner. Hvis man ser seg nødt til å samle inn individet, er det lurt å samle det i et glass med en bit tørkepapir og legge det i fryseren. Dyret kan da tas opp etter noen timer og prepareres pent for videre studier. Hvis det ikke er mulig å artsbestemme humlene på ytre karakterer, kan det være til hjelp å dissekere ut broddsliren på dronninger og arbeidere og genitaliene på hannene.

Det er ikke alltid like lett å artsbestemme humler i felt. Av og til kan det da være lurt å fange dem i et glass, slik at de kan studeres nærmere.
Referanser
Arnold SEJ, Idrovo MEP, Arias LJL, Belmain SR og Stevenson PC (2014). Herbivore defence compounds occur in pollen and reduce bumblebee colony fitness. J. Chem. Ecol. 40: 878–881.
Bengtson R (2011a). Inspirerende svensk bok om humler. (Bokanmeldelse av Humlor til Holmström, 2007). Fauna 64 (2–4): 106–108.
Bengtson R (2011b). Flott og lærerik svensk bok om humler. (Bokanmeldelse av Humlor i Sverige til Mossberg & Cederberg, 2012). Fauna 64 (2–4): 109–111.
Bengtson R (2013). Flott og god norsk humlebok, men med noen "krumspring". (Bokanmeldelse av Norges humler med Humleskolen til Bollingmo, 2012). Insekt-Nytt 38 (1): 29–34.
Bengtson R og Olsen KM (2013a). Lundgjøkhumle Bombus quadricolor i Norge. SABIMA-rapport. 40 s.
Bengtson R og Olsen KM (2013b). Kartlegging av rødlistede humler i Sør-Norge i 2011 og 2012. Kunnskapsstatus og forvaltning angående slåttehumle Bombus subterraneus, kløverhumle B. distinguendus, bakkehumle B. humilis, kysthumle B. muscorum, gresshumle B. ruderarius og lundgjøkhumle B. quadricolor. BioFokus-rapport 2013–2. 105 s.
Bengtson R og Olsen KM (2014a). Kartlegging av rødlistete humler sørøst i Norge i 2013. BioFokus-rapport 2014-1. 88 s.
Bengtson R og Olsen KM (2014b). Kartlegging av humler og dagsommerfugler i blomsterfelt på Steinssletta i Hole i Buskerud juli 2014. BioFokus-notat 2014-40. 36 s.
Benton T (2006). Bumblebees - The natural history and identification of the species found in Britain. Collins. 580 s.
Bollingmo T (1989). Worker – queen conflict and fitness consequences in a colony of Bombus hypnorum L. (Hymenoptera). Fauna Norv. Ser. B, 36: 69–73.
Bollingmo T (2012). Norges humler med Humleskolen. BRAINS Media. 295 s.
Cameron SA, Hines HM og Williams PH (2007). A comprehensive phylogeny of the bumble bees (Bombus). Biol. J. Linn. Soc. 91: 161–188.
Cameron SA, Lozier JD, Strange JP, Koch JB, Cordes N, Solter LF og Griswold TL (2011). Patterns of widespread decline in north American bumble bees. PNAS 108: 662–667.
Carolan JC, Murray TE, Fitzpatrick U, Crossley J, Schmidt H, Cederberg B, McNally L, Paxton RJ, Williams PH og Brown MJF (2012). Colour Patterns Do Not Diagnose Species: Quantitative Evaluation of a DNA Barcoded Cryptic Bumblebee Complex. Plos One 7: 662–667.
Cederberg B (1983). The role of trail pheromones in host selection by Psithyrus rupestris (Hymenoptera, Apidae). Ann. Ent. Fenn. 49: 11–16.
Cederberg B, Svensson BG, Bergström G, Appelgren M og Groth I (1984). Male marking pheromones in north European cuckoo bumble bees, Psithyrus (Hymenoptera, Apidae). Nova Acta Reg. Soc. Sci. Ups., Serie V:C, 3: 161–166.
Dafni A, Kevan P, Gross CL og Goka K (2010). Bombus terrestris, pollinator, invasive and pest: an assessment of problems associated with its widespread introductions for commercial purposes. Appl. Entomol. Zool. 45: 101–113.
Darvill B, Lepais O, Woodall LC og Goulson D (2012). Triploid bumblebees indicate a direct cost of inbreeding in fragmented populations. Molecular Ecology 21: 3988–3995.
Dramstad W (1996). Der humlene fremdeles surrer. FAGnytt Naturforvaltning 3 (4): 1–4.
Dramstad WE (1996). Do bumblebees (Hymenoptera, Apidae) really forage close to their nests? Journal of Insect Behavior 9: 163–182.
Dramstad W og Fry G (1995). Foraging activity of Bumblebees (Bombus) in relation to flower resources on arable land. Agric. Ecosyst. Environ. 53: 123–135.
Dupont YL, Damgaard C og Simonsen V (2011). Quantitative Historical Change in Bumblebee (Bombus spp.) Assemblages of Red Clover Fields. PLoS ONE 6 (9): e25172. DOI:10.1371/journal.pone.0025172
Dupont YL og Madsen HB (2010). Humlebier. Natur og Museum 49(1): 1–36.
Dupont YL, Strandberg B, Bruus M og Madsen HB (2015). Konkurrence mellem vilde bier og honningbier: Hvad ved vi egentlig? Tidsskrift for biavl 1/2015: 10–13.
Edwards M og Williams PH (2004). Where have all the bumblebees gone, and could they ever return? Brit. Wildl. June: 305–312.
Ellis JS, Knight ME, Darvill B og Goulson D (2006). Extremely low effective population sizes, genetic structuring and reduced genetic diversity in a threatened bumblebee species, Bombus sylvarum (Hymenoptera: Apidae). Molecular Ecology 15: 4375–4386.
Fjellstad W, Norderhaug A og Ødegaard F (2008). Tidligere og nåværende jordbruk. I Kålås, J.A., Henriksen, S., Skjelseth, S. & Viken, Å. (red.). Miljøforhold og påvirkninger på rødlistearter. Artsdatabanken, Norge. s. 39–49.
Forup ML og Memmott J (2005). The relationship between the abundances of bumblebees and honeybees in a native habitat. Ecological Entomology 30: 47–57.
Frankham R, Ballou JD og Briscoe DA (2002). Introduction to conservation genetics. Cambridge Univ. Press, Cambridge, UK. 617 s.
Free JB og Williams IH (1976). Pollination as a factor limiting the yield of field beans (Vivia faba L.). J. Agric. Sci. 87: 395–99.
Free JB og Ferguson AW (1986). Foraging of bees on oil-seed rape (Brassica napus L.) in relation to the flowering of the crop and pest control. J. Agric. Sci. 94: 151–54.
Fürst MA, McMahon DP, Osborne JL, Paxton RJ, Brown MJF (2014). Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature DOI: doi/10.1038/nature12977
Gallai N, Salles J-M, Settele J og Vaissière BE (2009). Economic valuation of the vulnerability of world agriculture confronted with pollinator. Ecol. Econ. 68: 810–821.
Gjershaug JO (2009). The social parasite bumblebee Bombus hyperboreus Schönherr, 1809 usurp nest of Bombus balteatus Dahlbom, 1832 (Hymenoptera, Apidae) in Norway. Norw. J. Entomol. 56: 28–31.
Gjershaug JO, Staverløkk A, Kleven O og Ødegaard F (2013). Species status of Bombus monticola Smith (Hymenoptera: Apidae) supported by DNA barcoding. Zootaxa 3716(3): 431–440.
Gjershaug JO og Ødegaard F (2012). Vurdering av risiko for biologisk mangfold ved innførsel av mørk jordhumle Bombus terrestris til Norge. NINA Rapport 895. 42 s.
Gjershaug JO, Ødegaard F og Staverløkk A (2012). Debattinnlegg: Kontroversiell ny bok om humler i Norge. Fauna 65 (1–2): 72–76.
Gjershaug JO, Ødegaard F, Staverløkk A og Dahle S (2013). Kartlegging av slåttehumle, kløverhumle, bakkehumle og lundgjøkhumle i 2013. NINA Rapport 997. 36 s.
González-Varo JP, Biesmeijer JC, Bommarco R, Potts SG, Schweiger O og Smith HG (2013). Combined effects of global change pressures on animal-mediated pollination. Trends in Ecology & Evolution 28(9): 524–30.
Goulson D (2003). Conserving wild bees for crop pollination. - Int. J. Food Agric. Environ. 1: 142–44.
Goulson D (2010). Bumblebees: behaviour, ecology and conservation. Oxford University Press. 317 s.
Goulson D (2013). A sting in the tale. Jonathan Cape, London. 265 s.
Goulson D (2015). Neonicotinoids impact bumblebee colony fitness in the field; a reanalysis of the UK’s Food & Environment Research Agency 2012 experiment. PeerJ 3:e854 https://dx.doi.org/10.7717/peerj.854.
Goulson D, Hanley ME, Darvill B, Ellis JS og Knight ME (2005). Causes of rarity in bumblebees. - Biol. Conserv. 122: 1–8.
Goulson D, Lye GC og Darvill B (2008). Decline and conservation of bumble bees. Annu. Rev. Entomol. 53: 191–208.
Goulson D, Whitehorn P og Fowley M (2012). Influence of urbanisation on the prevalence of protozoan parasites of bumblebees. Ecol. Entomol. 37: 83–89.
Greigsmith PW, Thompson HMN, Hardy AR, Bew MH, Findlay E og Stevenson JH (1994). Incidents of poisoning of honeybees (Apis mellifera) by agricultural pesticids in Great Britain, 1981-1991. Crop Prot. 13: 567–81.
Grixti JC, Wong LT, Cameron SA og Favret C (2009). Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 142: 75–84.
Holmström G (2007). Humlor – alla Sveriges arter. Så känner du igjen dem i naturen – och i trädgården. Brutus Östlings Bokförlag, Stockholm. 159 s.
Ings TC, Ward NL og Chittka L (2006). Can commersially imported bumble bees out-compete their native conspecifics? J. Appl. Ecol. 43: 940–948.
Kearns CA og Thomson JD (2001). The natural history of bumblebees, a sourchebook for investigations. University Press of Colorado, Boulder. 130 s.
Kosior A, Celary W, Olejnikzak P, Fijal J, Krol W, Solarz W og Plonka P (2007). The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of western and central Europe. Oryx 41: 79–88.
Kremen C, Williams NM og Thorp RW (2002). Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 99: 16812–16.
Kvisberglien E (2012). What determines the bumble bees' use of the roadside? Master's thesis in Environmental Health 2012. Faculty of Arts and Sciences, Telemark University College. 41 s.
Løken A (1973). Studies on Scandinavian bumble bees (Hymenoptera, Apidae). Norw. J. Entomol. 20: 1–218.
Løken A (1984). Scandinavian species of the genus Psithyrus Lepeletier (Hymenoptera: Apidae). Ent. Scand. Suppl. 23: 1–45.
Løken A (1985). Norske insekttabeller, 9. Humler. Norsk entomologisk forening, Ås. 39 s.
Madsen HB og Calabuig I (2012). Kommenteret checkliste over Danmarks bier – Del 5: Apidae (Hymenoptera, Apoidae). Ent. Medd. 80: 7–52.
MartinetB, Rasmont P, Cederberg B, Evrard D, Ødegaard F, Paukkunen P og Lecocq T (2015). Forward to the North: two Euro-Mediterranean bumblebee species now cross the Arctic Circle. Journal Annales de la Société Entomologique de France 51:303-309
McMahon DP, Fürst MA, Caspar J, Theodorou P, Brown MJF og Paxton RJ (2015). A sting in the spit: widespread cross-infection of multiple RNA viruses across wild and managed bees. Journal of Animal Ecology. DOI: 10.1111/1365-2656.12345
Meeus I, Brown MJF, de Graaf DC og Smagghe G (2011). Effects of invasive parasites on bumble bee declines. Conserv. Biol. 25: 662–671.
Meidell O (1934). Bier og humler i Rogaland (Apidae, Hym.). Stavanger Mus. Årsh. 43 (1932–33): 85–131.
Memmott J, Waser NM og Price MV (2004). Tolerance of pollination networks to species extinctions. Proc. R. Soc. London Sci. Ser. B 271: 2605–11.
Mjelde A (1981). Biologiske/økologiske sammenligninger av humleartene Bombus consobrinus Dahlbom og Bombus hortorum (L.) (Hymenoptera, Apidae). Hovedfagsoppgave i spesiell zoologi til matematisk-naturvitenskapelig embetseksamen ved Universitetet i Oslo 1981. 98 s.
Mjelde A (1983). The foraging strategy of Bombus consobrinus (Hymenoptera, Apidae). Acta Entomol. Fennica 42: 51–56.
Mossberg B og Cederberg B (2012). Humlor i Sverige. 40 artar att älska och förundras över. Bonnier Fakta. 192 s.
Nybø S, Skarpaas O, Framstad E og Kålås JA (2008). Naturindeks for Norge – forslag til rammeverk. NINA Rapport 347. 68 s.
Pekkarinen A (1979). Morphometric, colour and enzyme variation in bumblebees (Hymenoptera, Apidae, Bombus) in Fennoscandia and Denmark. Acta Zool. Fenn. 158: 1–60.
Pekkarinen A (1982). Morphology and specific status of Bombus lapponicus (Fabricius) and B. monticola Smith (Hymenoptera: Apidae). Ent. Scand. 13: 41–46.
Petterson MW, Cederberg B og Nilsson LA (2004). Grödor och vildbin i Sverige. Kunnskapssammenställning för hållbar utveckling av insektspollinerad matproduktion och biologisk mångfald i jordbrukslandskapet. Svenska Vildbiprojektet vid ArtDatabanken, SLU & Avdelningen för Växtekologi, Uppsala Universitet. 42 s.
Rasmont P (1984). Les bourdons du genre Bombus Latreille sensu stricto en Europe occidentale et centrale (Hymenoptera, Apidae). Spixiana 7: 135–160.
Pekkarinen A, Teräs I, Viramo J og Paatela J (1981). Distribution of bumblebees (Hymenoptera, Apidae: Bombus and Psithyrus) in eastern Fennoscandia. Not. Ent. 61: 71–89.
Rasmont P, Franzen M, Lecocq T, Harpke A, Roberts SPM, Biesmeijer K, Castro L, Cederberg B, Dvorák L, Fitzpatrick Ú, Haubruge E, Mahé G, Manino A, Michez D, Neumayer J, Ødegaard F, Paukkunen J, Pawlikowski T, Potts SG, Reemer M, Settele J, Straka J, Schweiger O (2015). Climatic Risk and Distribution Atlas of European Bumblebees. BioRisk 10, special issue, 234 pp. ISBN 978-954-642-768-7 (hardback) ISBN 978-954-642-769-4 (e-book)
Rasmont P og Mersch P (1988). Premiere estimation de la derive faunique chez les bourdons de la Belgique (Hymenoptera, Apidae). Ann. Soc. R. Zool. Belg. 118: 141–147.
Rundlöf M, Andersson GKS, Bommarco R, Fries F, Hederström V, Herbertsson L, Jonsson O, Klatt BK, Pedersen TR, Yourstone J og Smith HG (2015). Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature, DOI: 10.1038/nature14420.
Smissen J van der og Rasmont P (1999). Bombus semenoviellus Skorikov, 1910, eine für Westeuropa neue Hummelart (Hymenoptera: Bombus, Cullumanobombus). Bembix 13: 21–24.
Staverløkk A, Gjershaug JO og Ødegaard F (2012). Humler i Norge. En felthåndbok om våre 34 humlearter. NINA Temahefte 49. 26 s.
Svensson BG (1979). Pyrobombus lapponicus auct., in Europe recognized as two species: P. lapponicus (Fabricius, 1793) and P. monticola (Smith, 1849) (Hymenoptera, Apoidae, Bombinae). Ent. Scand. 10: 275–296.
Söderström B (2013). Sveriges humlor - en fälthandbok. Entomologiska föreningen i Stockholm. Malmö. 127 s.
Totland Ø, Hovstad KA, Ødegaard F og Åström J (2013). Kunnskapsstatus for insektpollinering i Norge – betydningen av det komplekse samspillet mellom planter og insekter. Artsdatabanken, Norge. 74 s.
Wermuth K og Dupont YL (2010). Effects of field characteristics on abundance of bumblebees (Bombus spp.) and seed yield in red clover fields. Apidologie 41:657–66.
Whitehorn PR, O'Connor S, Wackers FL og Goulson D (2012). Neonicotinoid pesticide reduces bumble bee colony growth and queen production, Science, 336 (6079), pp. 351-352.
Williams PH (1986). Environmental change and the distributions of British bumble bees (Bombus Latr.). Bee World 67: 50–61.
Williams PH (2005). Does specialization explain rarity and decline among British bumblebees? A response to Goulson et al. Biological Conservation 122: 33-43.
Williams PH (2012). Bombus – bumblebees of the world. – http://www.nhm.ac.uk/research-curation/research/projects/bombus/pr.html
Williams PH, Araujo MB og Rasmont P (2007). Can vulnerability among British bumblebee (Bombus) species be explained by niche position and breadth. Biological Conservation 138: 493-505.
Williams PH, Brown MJF, Carolan JC, An J-D, Goulson D, Aytekin AM, Best LR, Byvaltsev AM, Cederberg B, Dawson R, Huang J-X, Ito M, Monfared A, Raina RH, Schmid-Hempel P, Sheffield CS, Sima P og Xie Z-H (2012). Unveiling cryptic species of the bumblebee subgenus Bombus s. str. world-wide with COI barcodes (Hymenoptera: Apidae). Syst. Biodiv. 10: 21–56.
Williams PH, Byvaltsev AM, Cederberg B, Berezin MV, Ødegaard F, Rasmussen C, Richardson LL, Huang J, Sheffield CS og Williams ST (2015). Challenges to the Integrative Approach to Species Delimitation: Genes Suggest Arctic Bumblebees Combine Morphologically Cryptic Diversity with Ancestral Colour Polymorphisms that Cut Across Species. Innsendt manuskript.
Williams PH, Cameron SA, Hines HM, Cederberg B og Rasmont P (2008). A simpified subgeneric classification of the bumblebees (genus Bombus). Apidologie 39 (1): 46–74.
Williams PH og Osborne JL (2009). Bumblebee vulnerability and conservation world-wide. Apidologie 17: 1–21.
Öberg S, Gjershaug JO, Certain G og Ødegaard F (2010). Utvikling av metodikk for arealrepresentativ overvåking av utvalgte invertebratgrupper. Pilotprosjekt Naturindeks for Norge. NINA Rapport 555. 50 s.
Öberg S, Gjershaug JO, Diserud O og Ødegaard F (2011). Videreutvikling av metodikk for arealrepresentativ overvåking av dagsommerfugler og humler. Naturindeks for Norge. NINA Rapport 663. 56 s.
Öberg S, Pedersen B, Diserud OH, Gjershaug JO, Staverløkk A og Ødegaard F (2012). Dagsommerfugler og humler som tilstandsindikatorer i Naturindeks for Norge. Videre uttesting av metodikk og involvering av frivillige. NINA Rapport 836. 38 s.
Ødegaard F, Gjershaug JO og Mjelde A (2010). Forslag til norske navn på humler. Insekt-Nytt 35(1): 19–24.
Ødegaard F, Gjershaug JO, Staverløkk A og Mjelde A (2013). Faglig grunnlag for handlingsplan for kløverhumle Bombus distinguendus, slåttehumle Bombus subterraneus og bakkehumle Bombus humilis. NINA Rapport 816. 69 s.
Ødegaard F, Gjershaug JO, Öberg S og Mjelde A (2009). Status for humler (Hymenoptera, Apidae, Bombus spp.) i Norge 2010. Fauna 62: 94–104.
Aase AL, Ødegaard F, Mjelde A og Flagstad Ø (2011). Bombus subterraneus (L., 1758) (Hymenoptera, Apidae) rediscovered in Norway. Norw. J. of Ent. 58: 15–19.
Åström S, Åström J, Bøhn K, Gjershaug JO, Staverløkk A og Ødegaard F (2013). Dagsommerfugler og humler som tilstandsindikatorer i Naturindeks for Norge. Statusrapport etter årene 2009- 2013. NINA Rapport 1005. 66 s.
Åström S, Åström J, Bøhn K, Gjershaug JO, Staverløkk A og Ødegaard F (2014). Dagsommerfugler og humler som tilstandsindikatorer i Naturindeks for Norge. Oppsummering av året 2014. – NINA Rapport 1098. 26 s.