6SE Bioklimatiske seksjoner
Gradienten omfatter syv bioklimatiske seksjoner som fanger opp variasjon i humiditet og oseanitet på fastmark.
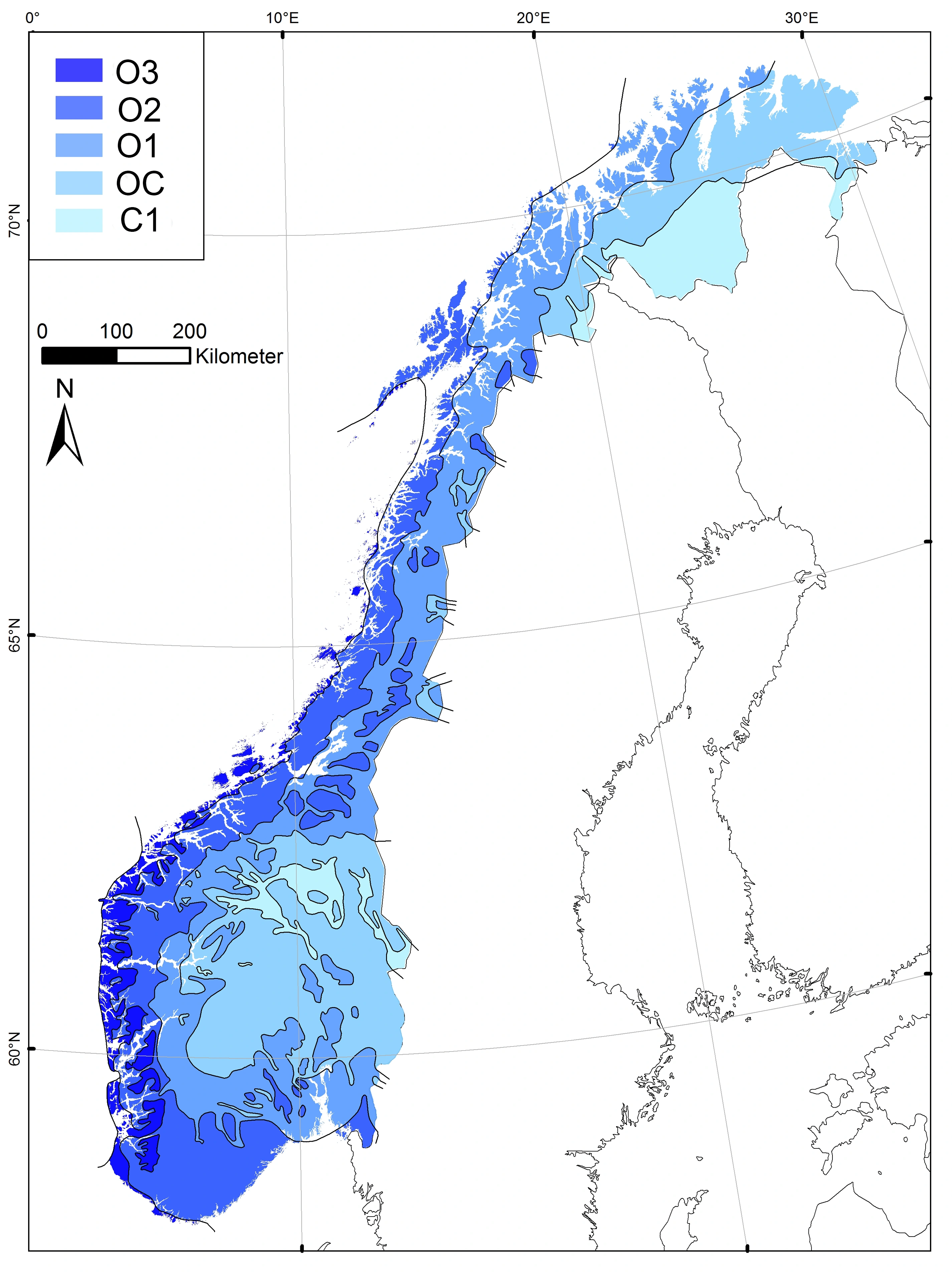
Alle organismer trenger vann (i tillegg til varme) for å opprettholde sine livsfunksjoner. Liksom varmekravet varierer også vannbehovet sterkt mellom arter og organismegrupper, fra arter som er tilpasset et helt liv i vann til landlevende arter som tåler sterk uttørking. Vanntilgangen varierer fra regional til svært fin lokal skala, og variasjonen i vanntilgang gjenspeiles i variasjon i arters utbredelse og forekomstmønstre over hele denne skalaspennvidden. På regional skala kommer variasjon i vanntilgang først og fremst til uttrykk som variasjon i humiditet. Humiditeten øker når nedbøren øker og når temperaturen avtar. Humid klima finnes i kystnære områder der fuktig havluft presses opp og avkjøles i møtet med høye fjell. Havet har imidlertid også en modererende effekt på temperaturklimaet. Kjølige somre og milde vintre, det vil si liten temperaturamplitude gjennom året (oseanisk klima), er typisk for kystnære områder, mens innlandet typisk har et tørt (kontinentalt) klima med varme somre og kalde vintre. Den regionale komplekse miljøvariabelen bioklimatiske seksjoner (6SE) fanger opp både humiditet og oseanitet. Bioklimatiske seksjoner (6SE) er en av de aller viktigste regionale komplekse miljøvariablene også på global skala, der spennvidden i vegetasjonsformasjoner fra regnskog via steppe til ørken fanges opp av sju bioklimatiske seksjoner. Av disse finnes alle bortsett fra den sterkest kontinentale seksjonen innenfor det området NiN dekker. Fem av seksjonene finnes på det norske fastlandet, den sterkest kontinentale bare på Spitsbergen. I Sør-Norge har den bioklimatiske seksjonsgradienten hovedretning fra vest til øst, mens den på Spitsbergen ikke har noen klar retning. På det norske fastlandet har arter med store krav til høy og stabil fuktighetstilgang et vestlig utbredelsesmønster, mens arter med stor toleranse overfor uttørking og/eller kalde vintre har et komplementært, østlig utbredelsesmønster.
Trinndeling
Trinninndelingen i seks trinn kalt seksjoner (sju på global basis) følger tradisjonen etter Ahti et al. (1968) for seksjonsinndeling av det norske fastlandet, med de tilpasninger som er gjort i den norske vegetasjonsseksjonsinndelingen, først av Dahl et al. (1986) og seinere av Moen (1998).
Moen (1998) følger Dahl et al. (1986) og utfigurerer en vintermild (termisk) underseksjon (O3t) innenfor sterkt oseanisk seksjon (O3) aller ytterst på vestlandskysten (se NiN[2]AR25: Fig. 3a). Denne underseksjonen karakteriseres ved forekomst av plantearter som for eksempel purpurlyng (Erica cinerea) og hinnebregne (Hymenophyllum wilsonii). Den andre (hygriske) underseksjonen, O3h, omfatter indre og høyereliggende strøk med et generelt fuktig (humid) klima. Todelingen av 6SE∙1 sterkt oseanisk seksjon (O3) videreføres ikke i NiN versjon 1 fordi Bakkestuen et al. (2008) viser at grensa mellom O3t og O3h ikke er basert på de samme kriteriene som de øvrige trinngrensene langs bioklimatiske seksjoner (6SE). Et skille mellom en termisk og en hygrisk del av den sterkt oseaniske seksjonen må derfor, om det skal opprettholdes, frikobles fra den bioklimatiske seksjonsgradienten. Variasjonen som ligger til grunn for å skille O3t fra O3h oppfattes imidlertid som så lite viktig (relevant for så få arter og naturtyper) at den ikke kommer til uttrykk i NiN versjon 2.
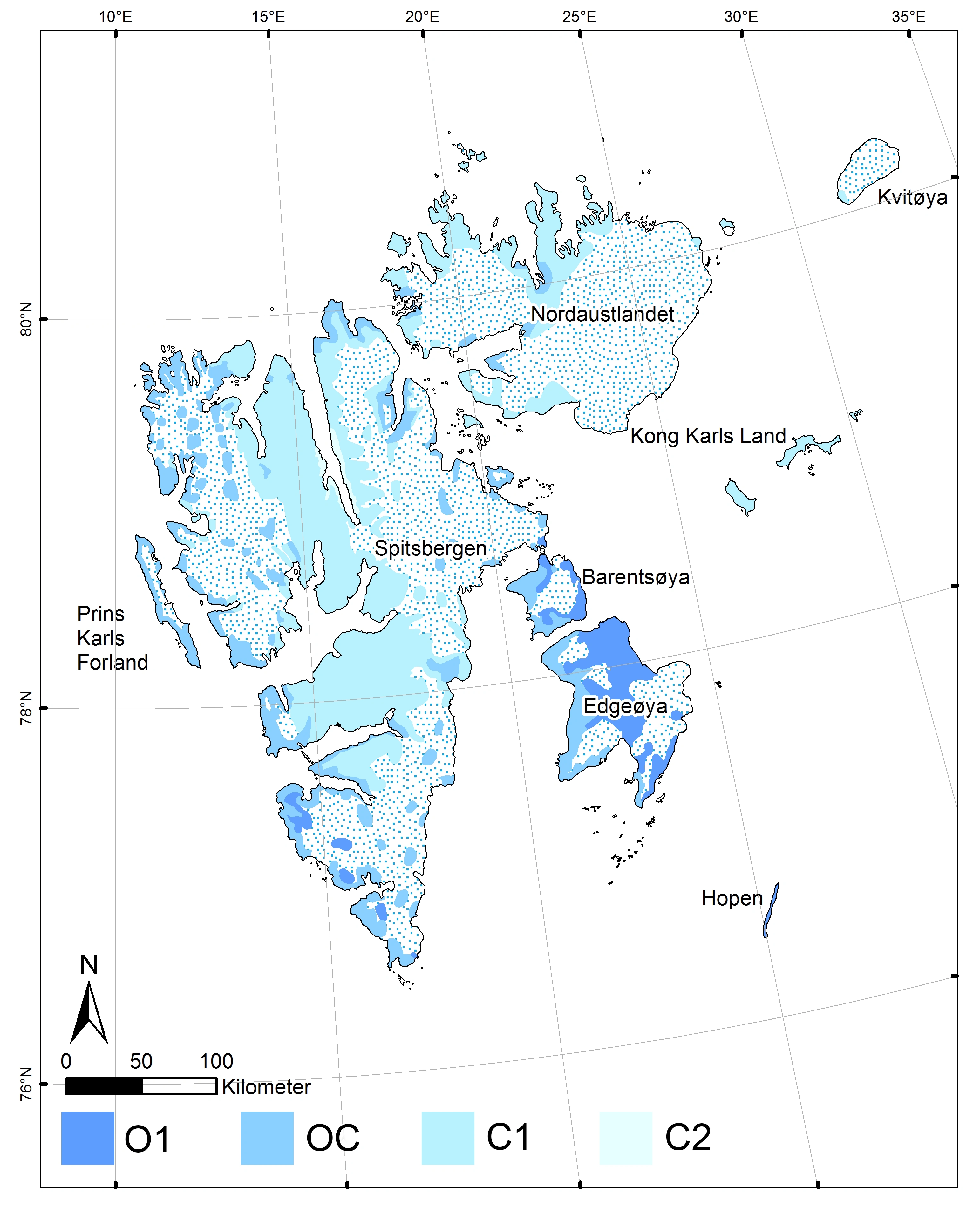
Seksjonsinndelingen av Svalbard foretatt på grunnlag av ekspertvurdering etter samme kriterier som inndelingen av bioklimatiske seksjoner (6SE) for det norske fastlandet (Moen 1998), er utarbeidet av Arve Elvebakk (se figuren til høyre).
Lokalt kan de regionale komplekse miljøgradientene bioklimatiske seksjoner (6SE) og bioklimatiske soner (6SO) variere sammen, ved at økende høyde over havet på samme tid innebærer overgang til en kjøligere bioklimatisk sone [høyere trinn langs bioklimatiske soner (6SO)] og til en mer humid bioklimatisk seksjon [lavere trinn langs bioklimatiske seksjoner (6SE)]. Årsaker til dette er at økende høyde over havet ikke bare innebærer redusert varmetilførsel, men også ofte økende nedbørmengder (Sjörs 1948, Førland 1979) som sammen med redusert fordampning (evapotranspi-rasjon) resulterer i økt humiditet. Ett eksempel på slike lokale sammenfall finnes i indre Østfold (R. Økland 1989b).
Trinnene langs bioklimatiske seksjoner (6SE) kan karakteriseres ved en kombinasjon av kriterier basert på målbare klimavariabler, hvorav de viktigste er vintertemperatur, nedbør, avstand fra kysten og snødekkevarighet [Bakkestuen et al. (2008); se også Tuhkanen (1980, 1984) og Moen (1998)]. Bioklimatiske seksjoner (6SE) er imidlertid ikke så sterkt relatert til hver enkelt av disse klimafaktorene som bioklimatiske soner (6SO) er til vekstsesonglengde og varmesum, og trinnene (seksjonene) er derfor vanskeligere å karakterisere klimatisk enn de bioklimatiske sonene. Humiditet (klimatisk fuktighet) er en funksjon både av temperatur og nedbør. Et mål på humiditet er nedbøroverskuddet, det vil si årsmiddelnedbøren minus evapotranspirasjonen (den samlete fordampningen fra en åpen vannflate i løpet av en tidsperiode, som i likhet med nedbøren måles i mm). Fordi evapotranspirasjonen avtar når temperaturen avtar, er humiditeten høyere på et sted med kjøligere klima enn på et sted med et varmere klima når nedbørmengden er den samme på de to stedene. Dette forklarer at et område på Spitsbergen som mottar 200 mm nedbør i året har et klima som kan karakteriseres som moderat humid [(og plasseres i overgangsseksjonen (OC)], mens et område i tropene som mottar 200 mm nedbør har et stort nedbørunderskudd og tilfredsstiller alle kriterier på ørken [«trinn 7» sterkt kontinental seksjon (C3), som ikke finnes innenfor NiN-området]. Tamms humiditetsindeks H (Tamm 1954) er en blant mange indekser for å angi nedbøroverskudd.
Bioklimatiske seksjoner (6SE) omfatter variasjon i, og relatert til, såvel humiditet som oseanitet, det vil si hygriske (fuktighetsrelaterte) og termiske (temperaturrelaterte) faktorer. Mangel på fullstendig samvariasjon mellom hygrisk (fuktighetsrelatert) og termisk (temperatur- og varmerelatert) variasjon gjør det vanskelig å finne et entydig sett av kriterier for trinninndeling. I NiN følger vi Moen (1998) og legger hovedvekten på hygriske kriterier, mens termiske kriterier (vintermildhet, sommertørke) blir tillagt mindre vekt.
Utfyllende karakteristikk av de seks trinnene, eller seksjonene, langs bioklimatiske seksjoner (6SE) for det norske fastlandet finnes i Moen (1998).
Kunnskapsbehov
Bioklimatisk seksjonsvariasjon kom i fokus i Norge før på slutten av 1980- og begynnelsen av 1990-tallet (Moen & Odland 1993). Seksjonsinndelingen i Moen (1998) er en konsensusinndeling basert først og fremst på botaniske kriterier. Studier, for eksempel av karplantearters utbredelse i Norge med multivariate metoder (ordinasjonsmetoder) framholder variasjonen som kommer til uttrykk gjennom bioklimatiske seksjoner (6SE) som den viktigste gradienten på regional skala, faktisk enda viktigere enn variasjonen som kommer til uttrykk gjennom bioklimatiske soner (6SO) (Pedersen 1990, Myklestad 1993). Ordinasjonsanalyse av relevante geologiske, topografiske og klimatiske variabler [«PCA-Norge»; Bakkestuen et al. (2008), se NiN[1]AR25: Fig. 1] viser at seksjonsgradienten fanger opp ett større spenn av miljøvariasjon enn bioklimatiske soner (6SO). Det er imidlertid effekten på den totale artssammensetningen som avgjør hvor viktig en RKM er for variasjon på natursystem-nivået, og i en totalvurdering anses sonegradienten derfor likevel å være viktigere enn seksjonsgradienten i Norge.
Fordi seksjonsbegrepet er relativt nytt i norsk biogeografisk sammenheng, finnes ingen samlet oversikt over arters eller naturtypers fordeling eller variasjonsmønstre langs seksjonsøkoklinen. Moen (1998) gir imidlertid fyldige karakteristikker av plantedekket i alle seksjoner på det norske fastlandet. På artsnivå kan informasjon om arters fordeling langs bioklimatiske seksjoner (6SE) til en viss grad trekkes ut av oversikter over arter med vestlig kontra østlig/nordøstlig utbredelsestyngdepunkt i Fennoskandia (jf. R. Økland 1989b, Dahl 1998). Påstanden om at kunnskapsmanglene om viktige detaljer i den regionale variasjonen i artssammensetning i norsk natur er store, særlig for andre organismegrupper enn karplanter [jf. beskrivelsen av bioklimatiske soner (6SO)], gjelder derfor også for bioklimatiske seksjoner (6SE). Spesifikke, viktige kunnskapsmangler kan oppsummeres i fem punkter:
- Det er behov for grundigere vurdering av hvorvidt 6SE∙6 klart kontinental seksjon (C2) finnes på det norske fastlandet (se NiN1)AR25).
- Til støtte for seksjonsinndelingen av Svalbard trengs en analyse av regional geoklimatisk variasjon på Svalbard etter mønster av «PCA-Norge» (Bakkestuen et al. 2008).
- Det er behov for en forbedret trinnløs seksjonsinndeling ved PCA-ordinasjon av miljøvariabler basert på et fullt sett av 1-km2 ruter (se NiN[1]AR25 og kommentar nedenfor). En slik analyse er under utarbeidelse (V. Bakkestuen et al., in prep.).
- Det er behov for mer kunnskap om regional variasjon i artssammensetning relatert til bioklimatiske seksjoner (6SE) for en rekke organismegrupper.
Det er behov for en grundig undersøkelse av hvordan lokaliteter med (og uten) egenskaper (arter og artssammensetning) typisk for «boreal regnskog» plasserer seg, både langs bioklimatiske soner (6SO) og langs bioklimatiske seksjoner (6SE). Dette kan for eksempel gjøres ved å se på plasseringen av lokaliteter med og uten slike kvaliteter langs trinnløse sone- og seksjonsmodeller (se NiN[1]AR25).
Kommentarer og tilleggsinformasjon
– Den første inndelingen av norsk Arktis i bioklimatiske seksjoner ble gjort i NiN versjon 1[seksjonsinndelingen av hele det sirkumarktiske området hos Elvebakk et al. (1999) er en generell biogeografisk inndeling og ikke en inndeling av den regionale komplekse miljøvariabelen bioklimatiske seksjoner (6SE) slik denne oppfattes i NiN].
– Trinnløs karakteristikk. En kort oppsummering av regionale komplekse miljøvariabler, identifisert ved hjelp av PCA-ordinasjon av geologiske, topografiske og klimatiske variabler og sammenlikning av denne med inndelingen i vegetasjonsseksjoner (Moen 1998), finnes i NiN[1]AR25. Fordi grunnlagsdata ikke var tilgjengelige for hele fastlands-Norge (bl.a. manglet meteorologiske data for km2-ruter der senterpunktet falt i hav) da den originale PCA-ordinasjonen ble gjort, vil en ny PCA-ordinasjon av alle ruter som inneholder land bli gjort i 2015 (V. Bakkestuen et al., in prep.). Denne vil bli lagt til grunn for å definere en ny trinnløs operasjonalisering av bioklimatiske seksjoner (6SE).
– Relevans for ferskvannssystemer. Bioklimatiske seksjoner (6SE) er relevant for beskrivelse av regional variasjon i fastmarkssystemer og våtmarkssystemer, men knapt i ferskvannssystemer. I sin oversikt over geografisk fordeling av arter fra en rekke artsgrupper i og ved ferskvann (inkludert karplanter, planteplankton, svamper, krepsdyr, insekter, bløtdyr og fisk), nevner J. Økland & K. Økland (1999) bare svært få arter med vestlig utbredelse, mens østlige utbredelsesmønstre først og fremst kan tilbakeføres til innvandrings- og spredningshistoriske forhold og ikke til bioklimatisk variasjon som sådan. Denne variasjonen fanges opp av RKM historisk ferskvannsforbindelse mot øst (6HF).
– Det finnes også variasjon i miljøforhold og artssammensetning på finere skala enn den regionale, som i noen grad er relatert til de samme miljøfaktorene som bioklimatiske seksjoner (6SE). Særlig gjelder dette de lokale komplekse miljøvariablene som uttrykker variasjon i tilgangen på vann som begrensende ressurs, og som er sterkt influert av det rådende nedbør- og temperaturklimaet på et gitt sted. De viktigste av disse er uttørkingsfare (UF) og uttørkingseksponering (UE). For begge disse øker arealdekningen av tørreste trinn mot kontinentale seksjoner og omvendt (arealdekningen av fuktigste trinn øker trinn mot oseaniske/humide seksjoner; Nordhagen 1943, R. Økland & Bendiksen 1985). Også arealdekningen av trinn langs LKM snødekkebetinget vekstsesongreduksjon (SV), oppfrysing (OF), rasutsatthet (RU) og vindutsatthet (VI) varierer systematisk langs bioklimatiske seksjoner (6SE). For alle disse LKM er regional variasjon i nedbørmengde, og dermed i snømengden, årsak til samvariasjon med den regionale gradienten. For snødekkebetinget vekstsesongreduksjon (SV) er det en direkte sammenheng ved at snømengden styrer arealdekningen av snødekning med ulik varighet. For oppfrysing (OF) er sammenhengen indirekte ved at kontinentale områder på grunn av lavere snømengder er sterkere utsatt for frostinntrengning i bakken enn oseaniske områder. Også for rasutsatthet (RU) er sammenhengen indirekte fordi ras- og skredhyppigheten generelt øker med økende snømengder.
– Begrepet «boreal regnskog» har vært gjenstand for mye diskusjon gjennom de siste 15–20 årene, blant annet i sammenheng med behov for forvaltningstiltak. Bakgrunnen for begrepet er at fastmarksskogsmark i områder med sterkt eller klart oseanisk klima (6SE∙1, O3, og 6SE∙2, O2) har flere karakteriserende fellestrekk (Holien & Tønsberg 1996), først og fremst en frodig epifyttisk lavflora og forekomst av enkeltarter (først og fremst lavarter) med en nordvestlig utbredelse i Norge. Forekomsten av slike typiske «boreale regnskogsarter» begrenser seg imidlertid ikke til fastmarksskogsmark; mange arter forekommer også hyppig blant annet i (T1) Nakent berg, grunntyper for bergvegg, og i (T13) Rasmark (G. Gaarder, pers. obs.). Dette indikerer at det ikke nødvendigvis er skogsmark i forhold til åpen mark som er utslagsgivende for den artssammensetningen som kjennetegner det «boreale regnskogsområdet», men heller at luftfuktigheten er stabil og så høy at uttørkingsfølsomme arter kan opprettholde stabile populasjoner over lang tid. Ekspertgruppa for NiN versjon 1 vurderte dette slik at norske forekomster av (T4) Fastmarksskogsmark med de egenskapene som kjennetegner «boreal regnskog» synes å fanges tilfredsstillende opp som variasjon langs en kombinasjon av de to regionale «økoklinene» bioklimatiske soner og bioklimatiske seksjoner, men erkjente samtidig at det er behov for grundigere analyse av hvordan lokaliteter med «regnskogskvaliteter» fordeler seg langs de to regionale «økoklinene». Et første steg i en slik analyse kan være å undersøke hvordan lokaliteter med og uten «regnskogsegenskaper fordeler seg langs trinnløse sone- og seksjonsmodeller» (se NiN[1]AR25).