M5 Afotisk marin sedimentbunn
Afotisk marin sedimentbunn omfatter natursystemer på ikke-stabiliserte sedimenter nedenfor kompensasjonsdypet, som ikke har naturlig permanent oksygenfrihet. Med ikke-stabilisert menes at sedimentene ikke er stabile nok til å ha stabile påvekstsamfunn.
Afotisk marin sedimentbunn omfatter variasjonen fra relativt beskyttete steder dominert av silt og leire til nokså eksponerte steder dominert av grus og småstein. Innholdet av organisk materiale varierer fra nesten helt uten organisk materiale (rein sand, grus og/eller stein) til nesten totaldominans av organiske sedimenter, og omfatter alle kornstørrelser fra den fineste leira til stein. Bunnfaunaen domineres av arter som lever nedgravd i sedimentet (infauna) og arter som lever på bunnen (epifauna). Mengden epifauna øker mot grovere og mer stabile sedimenter.
Stedvis kan faunaen domineres av større, bestandsdannende arter, f.eks. store hornkoraller (som først og fremst finnes i M2 Afotisk fast saltvannsbunn og på M6 Korallrev), eller, mer karakteristisk for sedimentbunn, av sjøfjær-arter eller svamper.
Mot større dyp finner det sted en gradvis utskifting av faunaen og en gradvis reduksjon i artsantallet for de fleste dyregrupper. Et viktig økologisk skille, som også viser seg i artssammensetningen, går omkring 700 meters dyp mellom atlantisk vann med temperaturer som varierer, men er over 0 °C, og dyphavsvann med stabile temperaturer under frysepunktet.

Afotisk marin sedimentbunn. Løs afotisk bunn med kontinuerlig oksygentilgang. Leirholdig bunn på ca. 350 m dyp (Ingøydypet utenfor Finnmark), med muddersjørose (Bolocera tuediae). Ei reke kan også ses i bildet.
Definisjonsgrunnlag og avgrensning
Det finnes ingen skarp, naturlig grense mellom M5 Afotisk marin sedimentbunn og M4 Eufotisk marin sedimentbunn. Grensa mellom hovedtypene er trukket ved kompensasjonsdypet, på samme dyp som grensa mellom M1 Eufotisk fast saltvannsbunn og M2 Afotisk fast saltvannsbunn, ved overgangen mellom dybdeintervallene der dybderelatert lyssvekking (DL) og dybderelatert miljøstabilisering (DM) avløser hverandre.
Variasjon
Variasjonen innenfor hovedtypen er systematisert ved bruk av S3-diagrammet, som er utførlig forklart i NiN[2] Artikkel 1, kapittel B3k. S3-diagrammet er delt opp i grunntyper på basis av diskusjoner i den marine faggruppa, og koordinert med inndelingen av M4. Som utgangspunktet for forenkling av S3-skjemaet mot økende dyp (basert på antakelsen om at det finner sted en homogenisering av artssammensetningen som følge av at leveforholdene blir mer stabile, men også mer ekstreme), er benyttet S3-skjemaet for M4 i det dypeste eufotiske beltet (DL∙d rødalgebeltet).
Denne hovedtypen omfatter også forekomster av organiske sedimenter; beskrevet som algegytjebunn (M4) i NiN versjon 1 (her: grunntyper for IO∙2). «Algegytjebunn» omfatter saltvannsbunn som regelmessig tilføres organiske sedimenter, blant annet alger, men også f.eks. maneter (Sweetman & Chapman 2011). Alger utgjør hovedmengden av det organiske materialet i den grunneste delen av dybdeintervallet som hovedtypen utspenner (DM∙1). Det er vist, blant annet fra Stillehavskysten utenfor California, at taredetritus kan samle seg i canyoner på havbunnen også på større dyp enn der alger kan leve (Vetter & Dayton 1999). Saltvannsbunn med høyt organisk innhold forekommer derfor også under kompensasjonsdypet. Organismesamfunnet på organisk bunn inneholder mange spesialiserte arter, for eksempel amfipoder. Liksom i deponier av organisk materiale på land (som for eksempel driftvoller) går nedbrytningen (’komposteringen’) relativt langsomt. «Algegytjebunnen» inneholder spesielle, dårlig kjente, men sannsynligvis artsrike organismesamfunn der blant annet flerbørstemark og krepsdyr er viktige. Hyppighet og utbredelse, blant annet mot større dyp, er ikke kjent.
De spesielle sorterte sedimentene (S3-klassene) skjellsand (S3–S∙A), svampspikelbunn (S3–S∙B) og korallgrus (S3–S∙C) er begrenset til dybdebeltene der organismene som gir opphav til dem, finnes i store nok mengder til å gi opphav til større, sammenhengende forekomster. Kolonier av store muslinger finnes bare i aller øverste del av det afotiske beltet (ned til kanskje 100 m), steinkorallrev mangler i dypvannsbeltet (under 600 m), mens kalk- og glassvamper kan finnes på alle dyp.
Svampspikelbunn (S3–S∙B) er sedimentbunn som for størstedelen utgjøres av svampspikler, det vil si rester etter svamper. Dyrerekka svamper (Porifera) omfatter tre klasser, kalksvamper (Calcarea), horn- og kiselsvamper (Demospongiae) og glassvamper (Hexactinidae), som produserer spikler av henholdsvis kalk, kisel og kitin, og silisium. Svamper er, i likhet med koralldyr, «filterspisere» som er avhengig av en viss strøm for å få tilfredsstilt sitt næringsbehov. Svampspikler er nålformete strukturer som fungerer som støtteskjelett for svampen. Svampspikelbunn arter seg derfor som en «glassvatt-matte». Svampspikler er bestandige. Der de forekommer i store mengder danner de en konsolidert sedimentbunn som er mye «luftigere» enn sedimentbunn som består av mineralmateriale. Svampspikelbunn kan inneholde oppløst oksygen helt ned til 1 m dyp! Faunasammensetningen på svampspikelbunn er dårlig utforsket, ikke minst fordi slike sedimenter er svært vanskelige å arbeide med! Det er grunn til å tro at de er like vanskelige å leve i! Det lever sannsynligvis mange arter i svampspikelsedimenter som er lite kjent, og det synes som om artssammensetningen på svampspikelbunn har et sterkt innslag av faunaelementer fra hardbunn. Det er ikke kjent for hvor mye spikler som må til for å få en artssammensetning som klart avviker fra artssammensetningen på «vanlig» bløtbunn, men sannsynligvis skjer det noe med artssammensetningen når spikeldominansen blir så stor at substratet blir oksygen-rikt og stabiliseres av svampspiklene. Grunntyper for svampspikelbunn (S3–S∙B) forekommer på steder som domineres av, eller har vært dominert av, svamper, eller i nærheten av svamp-dominerte steder, f.eks. under bergvegger med rikelig forekomst av svamper der døde svamp-deler drysser ned.
Mange svamparter er giftige og svamper spises av få andre dyr. Svamper huser en rekke ulike assosierte arter av andre dyregrupper og den assosierte artssammensetningen kan være særpreget. Svamper forekommer fra fjærebeltet ned til de største dyp, men svampdominerte natursystemer er kjent fra 30–40 m i fjordene (opp til rett under sprangsjiktet) og ned til 600–700 m sørpå og 1500 m nordpå (glassvamp-dominerte samfunn i «minusvann»). Svampdominert sedimentbunn har tentativt blitt beskrevet som egen hovedtype, «svampbunn», men det synes som om svampdominerte samfunn (liksom samfunn dominert av hornkoraller og sjøtrær) har en forutsigbar forekomst langs viktige LKM heller enn at de i seg sjøl virker sterkt strukturerende på artssammensetningen. Dominans av svamper, hornkoraller eller annen stasjonær megafauna skal beskrives ved hjelp av beskrivelsessystemets variabel dekning av stasjonær megafauna (1AG–H), eventuelt med spesifisering av dominerende artsgruppe relativ sammensetning av fastsittende megafauna (1AR–H–X) der X indikerer hvilken del-artsgruppe som dominerer.
Korallgrusbunn (S3–S∙C) har samme relasjon til M6 Korallrev som svampspikelbunn har til svampdominert sedimentbunn, og omfatter lokaliteter som er preget av nærhet til eller tidligere forekomst av korallrev.
Konsolidert leire, eller moreneleire (S3–E∙4 × S3–F∙3), er kjent fra alle dyp (M. Dolan, pers. medd.).
Det er ikke klart hvor mange standardtrinn variasjonen i artssammensetning gir grunnlag for å dele DM i- Bortsett fra det tydelige skillet omkring grensa mellom atlantisk vann og dyphavsvann ved ca. 700 m (varierer litt fra område til område), skjer endringene gradvis og omfanget av endringer varierer sikkert fra gruppe til gruppe. Høisæter (2010) beskriver variasjonen i artssammensetning for snegler i Norskehavet, og finner endring i artssammensetning langs hele dybdegradienten, men sannsynligvis ikke mer enn til å gi grunnlag for 3 standardtrinn. Femtrinnsoppdelingen av DM i NiN 2 kan derfor vise seg for fin.
På sandbunn (sannsynligvis også andre substrater) finnes dokumentert variasjon i artssammensetning som sannsynligvis er relatert til variasjon i strømforhold; f.eks. synes det som om dominans av sjøfjær er typisk for litt mer strømutsatte steder. Det er uklart hvor stor variasjon i artssammensetning det er tale om, og VF er derfor tentativt inkludert som uLKM i beskrivelsessystemet for M5.
Sjøfjær-arter (Pennatulacea), bl.a. vanlig sjøfjær (Pennatula phosphorea), bambuskorall (Isidella lofotensis) som er en slektning av sjøbusk og sjøtre, og andre hornkorall-arter skiller, seg fra de «skogdannende» hornkorall-artene [sjøtre (Paragorgia arborea), men også sjøbusk (Paramuricea placomus) og risengrynkorall (Primnoa resedaeformis)] ved å ha en fotliknende utvidelse i nedre del, som forankrer dem i sedimentbunn. Disse artene danner mindre tette kolonier og er mindre sterkt strukturerende på den øvrige artssammensetningen enn fastbunnshornkorallene, bunn der de dominerer inngår i M5 Afotisk marin sedimentbunn.
DD er tentativt inkludert som uLKM for å gi mulighet for å beskrive den spesielle dyphavsfaunaen i dype terskelfjorder som f.eks. Sognefjorden og Tysfjord.
Kunnskapsbehov
– Det er sterkt behov for sammenstilling av generaliserte artslister for hovedtypen, både med sikte på å evaluere graden av oppdeling av S3-diagrammet og antallet trinn som DM deles opp i.
– Det er behov for grundigere analyser av hvorvidt svamp- og hornkoralldominert sedimentbunn faktisk inneholder en sterkt avvikende artssammensetning, betinget av fastsittende megafauna som strukturerende artsgruppe. Inntil dette spørsmålet er avklart, blir verken «svampbunn’ eller «korallskog-bunn’ akseptert som hovedtyper i NiN.
Grunntypeinndeling
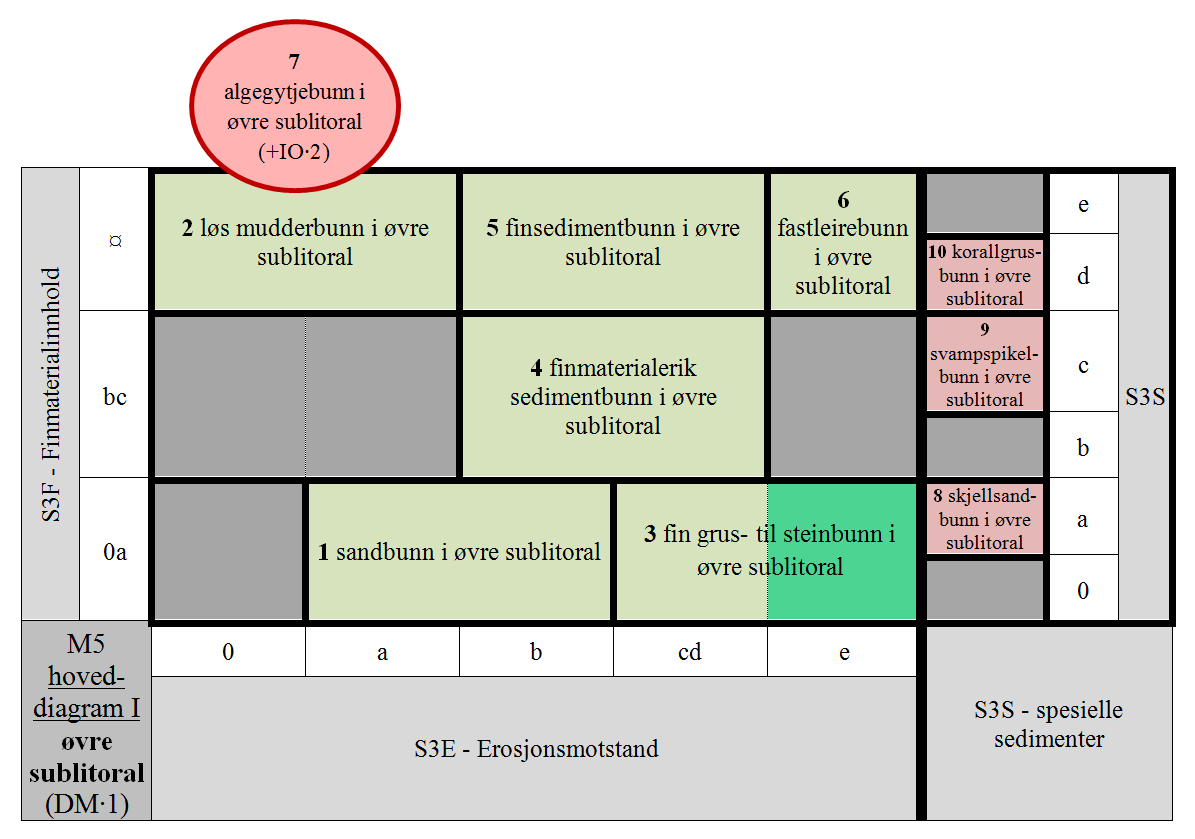
M5 er delt i 36 GT for antatt realiserte kombinasjoner av hLKM S3 og DM, hvorav 8 for S3–S, + 2 GT for spesielle kombinasjoner av hLKM S3 og DM, for IO∙2.
- M5-1 uten finmateriale og finmaterialfattig afotisk bunn med liten erosjonsmotstand i øvre sublitoral
- M5-2 finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i øvre sublitoral
- M5-3 uten finmateriale og finmaterialfattig afotisk bunn med intermediær til stor erosjonsmotstand i øvre sublitoral
- M5-4 litt og temmelig finmaterialrik afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i øvre sublitoral
- M5-5 finmaterialdominert afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i øvre sublitoral
- M5-6 finmaterialdominert afotisk bunn med stor erosjonsmotstand i øvre sublitoral
- M5-7 organisk finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i øvre sublitoral
- M5-8 afotisk skjellsandbunn i øvre sublitoral
- M5-9 afotisk svampspikelbunn i øvre sublitoral
- M5-10 afotisk korallgrusbunn i øvre sublitoral
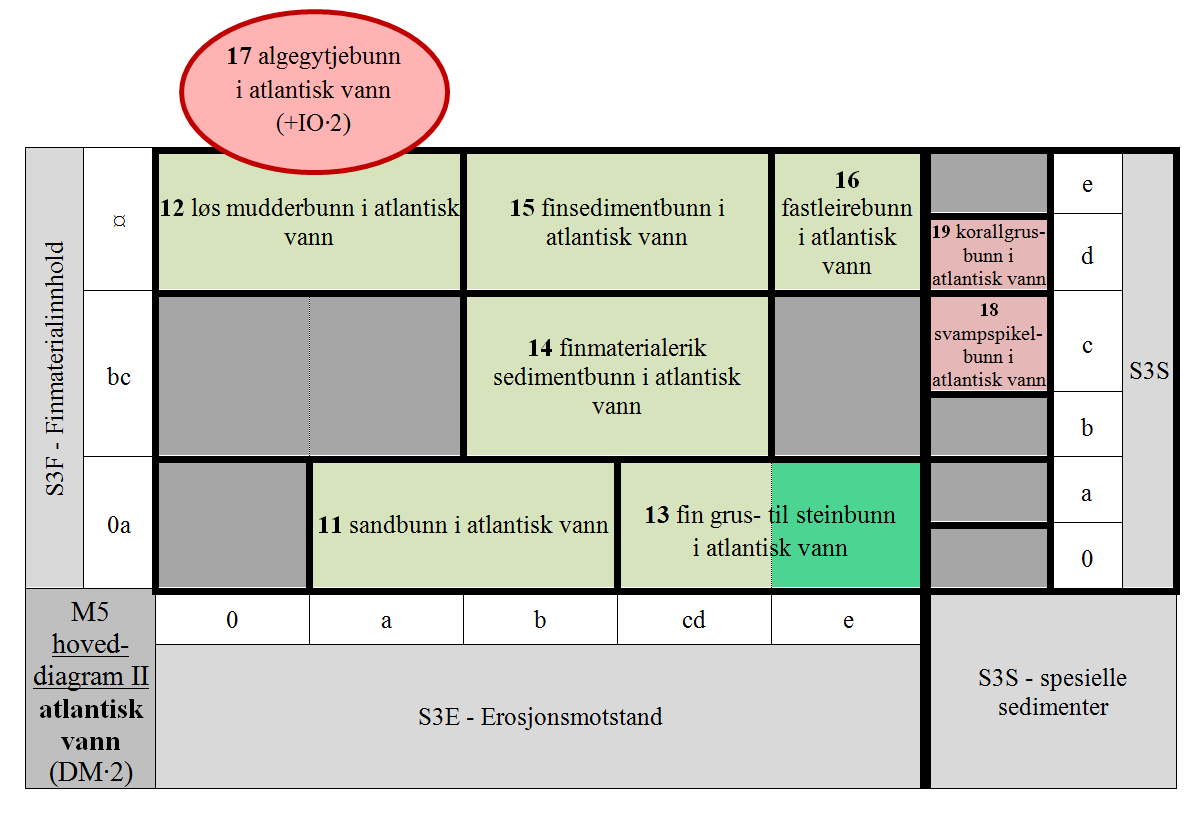
- M5-11 uten finmateriale og finmaterialfattig afotisk bunn med liten erosjonsmotstand i atlantisk vann
- M5-12 finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i atlantisk vann
- M5-13 uten finmateriale og finmaterialfattig afotisk bunn med intermediær til stor erosjonsmotstand i atlantisk vann
- M5-14 litt og temmelig finmaterialrik afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i atlantisk vann
- M5-15 finmaterialdominert afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i atlantisk vann
- M5-16 finmaterialdominert afotisk bunn med stor erosjonsmotstand i atlantisk vann
- M5-17 organisk finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i atlantisk vann
- M5-18 svampspikelbunn i atlantisk vann
- M5-19 korallgrusbunn i atlantisk vann
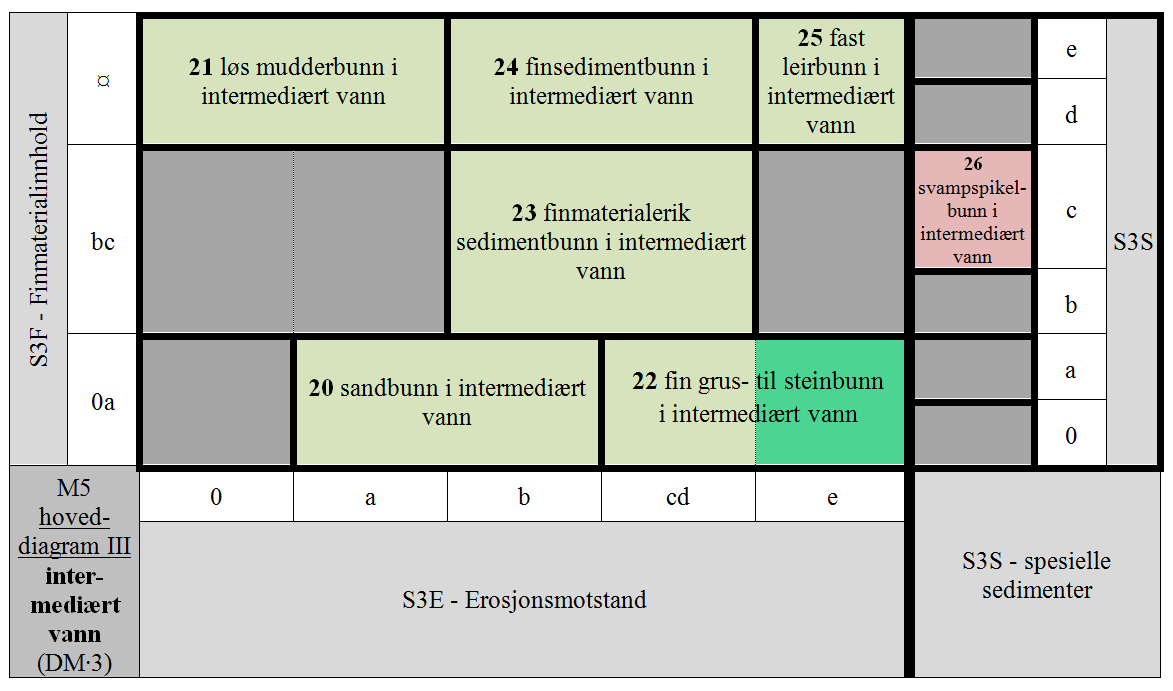
- M5-20 uten finmateriale og finmaterialfattig afotisk bunn med liten erosjonsmotstand i intermediært vann
- M5-21 finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i intermediært vann
- M5-22 uten finmateriale og finmaterialfattig afotisk bunn med intermediær til stor erosjonsmotstand i intermediært vann
- M5-23 litt og temmelig finmaterialrik afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i intermediært vann
- M5-24 finmaterialdominert afotisk bunn med temmelig liten til temmelig stor erosjonsmotstand i intermediært vann
- M5-25 finmaterialdominert afotisk bunn med stor erosjonsmotstand i intermediært vann
- M5-26 svampspikelbunn i intermediært vann
- M5-27 uten finmateriale og finmaterialfattig afotisk bunn med liten erosjonsmotstand i bathyal
- M5-28 finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i bathyal
- M5-29 uten finmateriale og finmaterialfattig afotisk bunn med intermediær til stor erosjonsmotstand i bathyal
- M5-30 finmaterialdominert bunn med temmelig liten til temmelig stor erosjonsmotstand i bathyal
- M5-31 finmaterialdominert bunn med stor erosjonsmotstand i bathyal
- M5-32 svampspikelbunn i bathyal
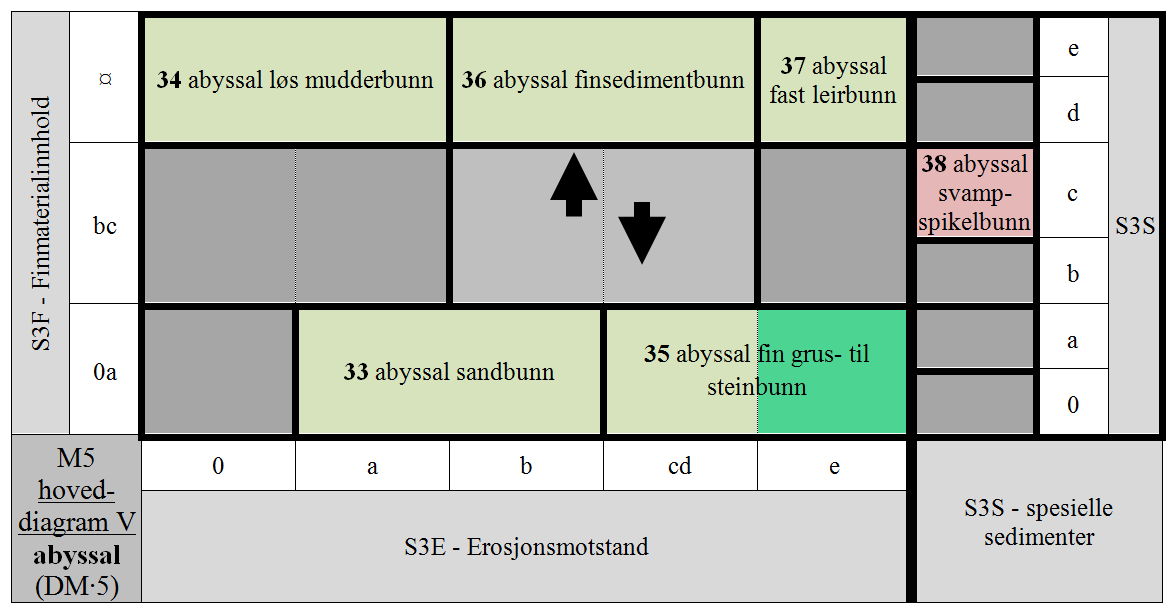
- M5-33 uten finmateriale og finmaterialfattig afotisk bunn med liten erosjonsmotstand i abyssal
- M5-34 finmaterialdominert afotisk bunn med ingen og svært liten erosjonsmotstand i abyssal
- M5-35 uten finmateriale og finmaterialfattig afotisk bunn med intermediær til stor erosjonsmotstand i abyssal
- M5-36 finmaterialdominert bunn med temmelig liten til temmelig stor erosjonsmotstand i abyssal
- M5-37 finmaterialdominert bunn med stor erosjonsmotstand i abyssal
- M5-38 svampspikelbunn i abyssal