Tenthredininae


Underfamilien Tenthredininae består av middels til store, til dels langstrakte og fargerike arter. Rundt 2000 arter er kjent på verdensbasis. Over 200 arter forekommer i Europa. 56 av disse er kjent fra Norge.
- Innhold
- Karakteristika og taksonomi
- Egg og larve
- Vertsplanter og ernæring
- Formering
- Det voksne stadiet
- Flygetid og livslengde
- Spredningsevne
- Habitat
- Innsamling av planteveps
Karakteristika og taksonomi
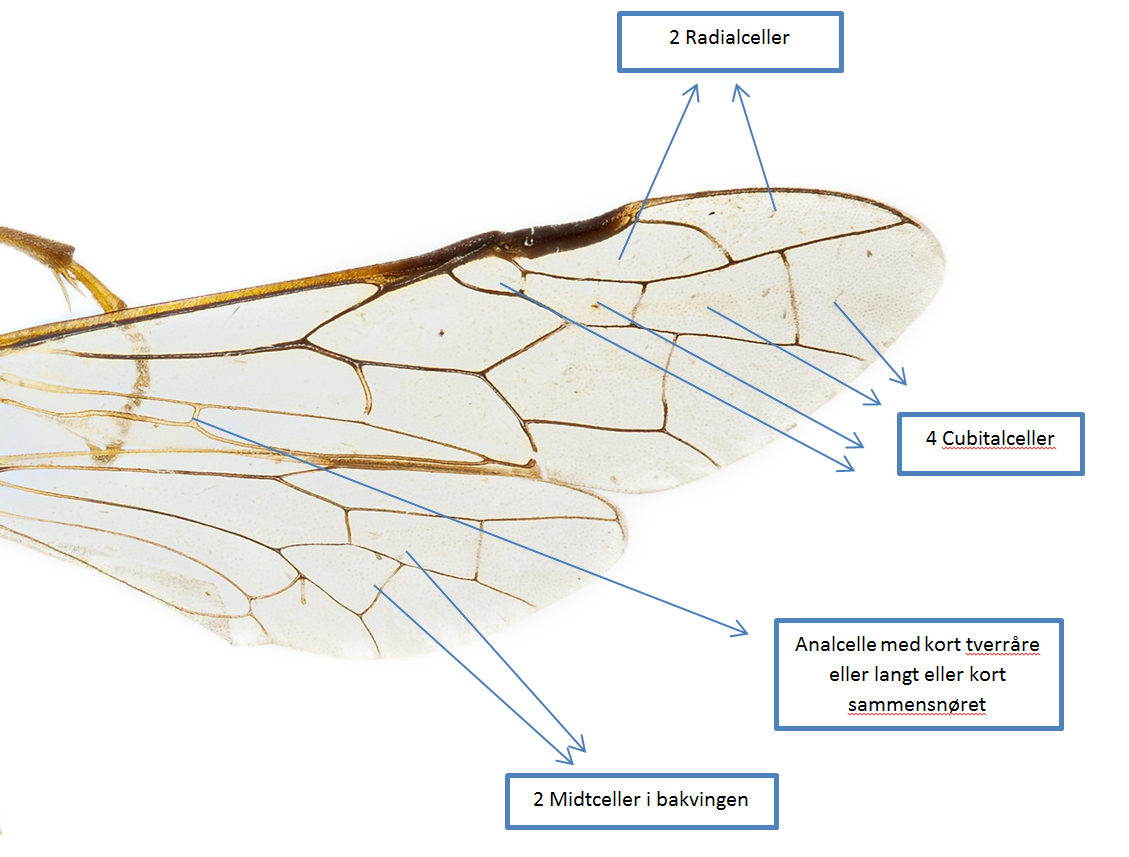
Figur 1
Generelle karakterer som i kombinasjon kan brukes til identifisering av Tenthredininae. Bildet viser vingene til en Tenthredo-art.
Karakterer på vingene er kanskje det som lettest skiller denne underfamilen fra andre underfamilier innen Tenthredinidae. Følgende kombinasjon av vingekarakterer kan benyttes for identifisering av europeiske arter innen Tenthredininae (figur 1):
- 2 Radialceller.
- 4 Cubitalceller.
- Analcellen er enten med kort tverråre eller langt eller kort sammensnøret.
-
Bakvingen har 2 midtceller (hannene til Perineura mangler derimot midtceller, og hos hannene til både Perineura og Tenthredopsis finnes i tillegg en ytre randåre i bakvingen).
Karakter 1 og 2 forekommer også innen underfamiliene Blennocampinae, Allantinae, Selandriinae og hos enkelte Nematinae. Hos Blennocampinae passer derimot ikke karakter 4. Hos mange Allantinae passer ikke karakter 2 (disse har ofte bare tre Cubitalceller). Hvis karakter 2 allikevel er til stede, vil ikke karakter 3 og 4 passe for disse. Hos mange Selandriinae passer karakter 1, 2 og 4, mens karakter 3 er annerledes. Hos Nematinae passer som regel ikke karakter 1 for de fleste slektene (de fleste Nematinae har ingen tverråre som deler radialcellen i to). Har de derimot dette, vil karakter 3 ikke passe. Et unntak er slekten Hoplocampa. Artene i denne slekten har lignende årenett i vingene som Tenthredininae, men er ellers veldig ulike. Denne lille slekten utgjøres av til dels svært små svarte eller gule arter som flyr tidlig på våren. Hoplocampa er knyttet til ulike vedaktige planter innen rosefamilien (Rosaceae).
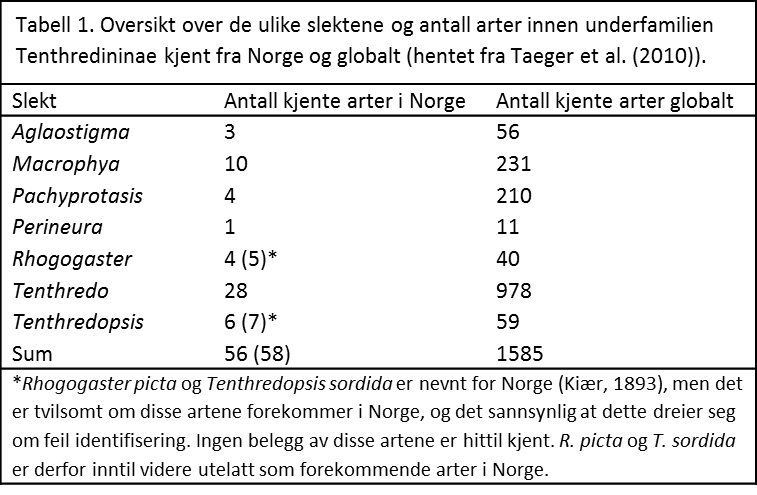
Underfamilien er særlig artsrik i Asia, spesielt i Kina og tilgrensende områder. På verdensbasis er det kjent nesten 2000 arter (Taeger et al. 2010), noe som gjør dem til den mest artsrike gruppen med planteveps. 56 av disse artene er kjent fra Norge. Underfamilien består av mange slekter, men spesielt slekten Tenthredo er svært artsrik med nærmere 1000 kjente arter på verdensbasis (Taeger et al. 2010). I Norge er følgende slekter representert: Aglaostigma, Macrophya, Pachyprotasis, Perineura, Tenthredo og Tenthredopsis. Tabell 1 gir en oversikt over hvor mange arter som er kjent fra Norge i hver slekt. Mange av disse er utbredt over store deler av landet og er vanlige, men mange har også en mer sørlig utbredelse, og er bare kjent fra Sør-Norge. Enkelte har også en begrenset utbredelse, og forekommer bare i de varmeste delene av Sør-Norge, som Oslofjord-regionen og langs Sørlandskysten, eller bare i Nord-Norge. Noen få arter er også svært sjeldne.
I Europa forekommer ytterligere noen slekter, Sciapteryx, Siobla og Ussurinus, men disse er ennå ikke funnet i Norge. Selv om man langt på vei har kunnskap om hvilke arter som forekommer i Norge, kan man nok forvente nye arter innen Tenthredininae for Norge i fremtiden, kanskje særlig på grunn av splitting av arter som følge av revisjoner, men også at alle arter som er her trolig ennå ikke er funnet. Enkelte arter som forekommer i Sør-Sverige, men ikke i Norge, kan nok også tenkes å etablere seg i Norge før eller siden.
-


- Lisens
- CC BY 4.0
- Opphav
- Ole Jørgen Lønnve
- Utgiver
- Biofokus
- Last ned (23.7 KB)
- Rediger (Krever innlogging)
Egg og larve
Eggleggingen skjer ved at hunnen lager et snitt i plantevevet hvorpå egget eller eggene legges. Eggskallet er tynt og elastisk. Eggleggingsorganet til hunnen er formet som en sag, derav navnet «sawflies» på engelsk. Sagens og tennenes fasong og antall er viktige karakterer for riktig identifisering av mange arter. Larvene minner om sommerfugllarver og spiser bladverket til vertsplanten. De har tre par gangføtter og åtte par vorteføtter. Når larven er ferdig utvokst, graver den seg ned i substratet på bakken hvorpå den spinner en kokong. Overvintringen skjer som prepuppe inne i kokongen.

Vertsplanter og ernæring

I motsetning til arter innen de fleste andre plantevepsgrupper, er mange av artene innen Tenthredininae svært polyfage, dvs at de går på en lang rekke til dels svært ulike vertsplanter (Gould & Bolton 1988). F. eks. er Tenthredo ferruginea beskrevet å gå på vidt forskjellige vertsplanter som einstape og rogn (Taeger et al. 1998). Enkelte arter går også på gress og starr. Derimot går ingen arter på bartrær. Vertsplantevalget er derimot ikke tilstrekkelig klarlagt for en rekke arter.
Det voksne stadiet til mange planteveps er, i motsetning til larvestadiet, ofte rovdyr som tar andre insekter. Særlig mange arter innen Tenthredo og spesielt Rhogogaster er glupske rovdyr som spiser mindre insekter som tovinger. Enkelte kan også spise sine artsfrender (Viitasaari 2002).
Mange arter søker til blomster, særlig arter innen Tenthredo, spesielt arter i Tenthredo arcuata-komplekset, men også arter i Rhogogaster er hyppig å finne på blomster. Skjermplanter (Apiaceae), som sløke, kvann, hundekjeks og skvallerkål, blir besøkt av en rekke arter, men også seljerakler, engsoleie og ryllik samt en rekke andre blomsterplanter blir besøkt (figur 3 og 4). Generelt oppsøker planteveps blomster for å: 1) spise pollen og nektar, 2) finne eller vente på et bytte (mindre insekter) de kan spise eller 3) finne seg en make (Liston 1980). Selv om dette er lite studert hos planteveps, er det sannsynlig at mange arter spiller en viktig rolle for pollineringen av enkelte blomsterplanter (Goulet 1996, Totland et al. 2013). Dette gjelder særlig arter innen Tenthredo arcuata-komplekset. Ofte kan man finne individer innen dette komplekset som har store mengder pollen på kroppen. Plantevepsene flyr fra blomst til blomst, og det er derfor nærliggende å anta at de også kan spille en rolle for pollinering. I tillegg kommer at artene innen dette komplekset ofte er svært tallrike. Utenfor Norge, spesielt i Sør- og Mellom-Europa og videre østover, forekommer en gruppe Tenthredo-arter (underslekten Elinora) med spesielt lange munndeler tilpasset pollenspising (Liston 1980, Blank & Taeger 2006). Artene i denne gruppen går på korsblomster (Brassicaceae), og de voksne oppsøker blomstene til disse.

Formering
De fleste artene innen denne underfamilien har kjønnet formering, dvs at hanner er kjent for de fleste artene innenfor den globale utbredelsen til arten. Det betyr derimot ikke at alle artene formerer seg kjønnet innenfor hele deres utbredelsesområde. Hos oss finnes flere eksempler på arter der hanner ennå ikke er kjent (f.eks. Macrophya teutona og Aglaostigma gibbosum) eller der hanner er mindre vanlige enn hunnene (f.eks. Tenthredo atra). Slike arter formerer seg derfor helt eller delvis partenogenetisk hos oss. Ukjønnet formering er derimot vanligere innenfor andre grupper planteveps enn hos Tenthredininae (f.eks. innenfor underfamiliene Blennocapinae og Nematinae). Hos de fleste plantevepser vil befruktete egg utvikles til hunner, mens ubefruktete egg utvikles til hanner.
Det voksne stadiet

Figur 5
MangeTenthredo-arter kan minne om broddveps (Aculeata) i farger. Ofte kan man også finne Tenthredo-arter side om side med ulike arter broddveps eksponert på blomstrende skjermplanter. Bildet viser bladvepsen Tenthredo vespa og gravevepsen (Crabronidae) Mellinus arvensis.
Hos mange av artene innen Tenthredininae er det klare visuelle forskjeller mellom kjønnene. Forskjellene går som regel på at kjønnene har ulike farger på deler av kroppen, f.eks. ben eller bakkropp. Generelt er hannene også noe mindre og tynnere enn hunnene, men de er som regel mer aktive og flyr mer rundt. Kjønnsforskjeller i farger finner vi særlig innen slektene Tenthredopsis og hos mange Tenthredo- og Macrophya-arter. Hos andre slekter, spesielt hos Rhogogaster, men også hos mange Tenthredo-arter, er derimot kjønnsforskjellene små. Enkelte arter varierer også en del i farger. Dette kan være geografisk betinget, eller være mer generelt. F.eks. varierer Tenthredo livida-hunnene ganske mye. På Østlandet er de som regel helt svarte, mens det spesielt på Vestlandet og i Nord-Norge forekommer hyppig individer med rødt på bakkroppen. Former med rødt på bakkroppen er også kjent fra andre steder, f. eks. i Alpene. Enkelte arter viser også en enda større variasjon hvis vi beveger oss utenfor Norden. Tenthredopsis litterata er eksempel på en slik art. I Norge forekommer to former, én der hunnens bakkropp er helt svart og én annen der bakkroppsspissen er rød. Sørover i Europa forekommer derimot andre fargevarianter av denne arten (Blank et al. 1998).
Mange arter minner i habitus om andre veps, spesielt enkelte broddveps (figur 5). Dette fenomenet kalles mimikry. Flere arter innen slekten Macrophya likner på veiveps (Pompilidae) både i kropp og til dels i adferd, mens arter innen Tenthredo-arcuata-komplekset kan minne om bier. Noen arter kan også i felt minne om store parasittveps (Ichneumonidae), f.eks. Tenthredo livida og T. mandibularis. Å etterligne andre veps kan være lurt. Både veiveps, graveveps, bier og enkelte parasittveps kan, i motsetning til planteveps, stikke.
Enkelte arter i innen andre bladvepsgrupper kan ved første øyekast også minne om Tenthredininae. F. eks. kan Allantus togatus (Allantinae) (figur 6) i ytre trekk likne en del på f.eks. Tenthredo vespa eller T. neobesa.

{kind=link}
Figur 6
Allantus togatus hunn. Denne arten kan ved første øyekast minne om enkelte Tenthredo-arter. Årenettet på vingene er derimot annerledes.
Flygetid og livslengde
De fleste artene har hos oss én generasjon i året. Det voksne stadiet er hos mange arter kortlevd, men enkelte arter kan nok leve en stund. Dette gjelder i hovedsak de artene som tar til seg næring som voksne, og særlig arter innen slektene Tenthredo og Rhogogaster.
Flygetiden hos Tenthredininae starter hos de fleste artene ikke før på forsommeren. I Sør-Norge som regel fra slutten av mai og utover i juni. I Nord-Norge og i fjellet som regel senere. Tenthredininae er derfor noe senere på vingene enn mange arter innen andre grupper planteveps, f.eks. mange arter innen Nematinae, Selandriinae og Blennocampinae, som kan ha en svært tidlig flygetid. F. eks. kan enkelte Dolerus-arter (Selandriinae) være på vingene allerede i slutten av mars. Derimot flyr mange Tenthredininae i en periode da mange andre planteveps er ferdig med sin flygetid. Enkelte arter flyr også utover i juli og inn i august. Dette gjelder særlig arter innen Tenthredo arcuata-komplekset og Tenthredo mesomalas-gruppen. Hos spesielt Tenthredo arcuata-komplekset er det sannsynlig at det kan være mer enn én generasjon per sesong hos oss. Enkelte andre arter kan nok under spesielle forhold en sommer ha mer enn én generasjon, men dette er ikke vanlig.
Spredningsevne
Planteveps generelt er antatt å ha en forholdsvis dårlig spredingssevne (Viitasaari 2002). De er ikke spesielt gode flygere. Dette gjelder også underfamilien Tenthredininae. Således vil som regel artsantallet av planteveps på øyer og andre isolerte steder være lavere enn på fastlandet. Fra Norge er det ikke gjort detaljerte studier som belyser dette godt nok, men studier gjort på øyene i indre Oslofjord, indikerer at artsantallet kan være noe lavere her enn bare noen kilometer unna på fastlandet (Endrestøl et al. 2007).
Habitat
Underfamilien er representert i de fleste habitater hos oss, men det er til dels stor forskjell på hvilke habitater ulike arter foretrekker. Arter særlig i slektene Tenthredopsis og Aglaostigma samt flere Tenthredo-arter foretrekker eng- og kanthabitater (figur 7). Mange Tenthredo- og Macrophya-arter foretrekker skog med rikt innslag av løv og frodig undervegetasjon, mens enkelte andre arter, spesielt Rhogogaster, er særlig tallrike i fjellbjørkeskogen og i vierbeltet i fjellet (figur 8). Noen arter foretrekker spesielt varme lokaliteter (f.eks. Tenthredo scrophulariae,Tenthredopsis litterata og Macrophya montana), mens Tenthredo moniliata og Aglaostigma gibbosum foretrekker mer fuktige og sumpaktige habitater. Kanter mot vassdrag, hvor det er en mosaikk av løvskog med innslag av frodig, fuktig og kraftig vegetasjon med diverse gress-, starr-, bregne-arter og forskjellige høystauder, er også habitat for en rekke arter, f. eks. kan de litt uvanlige artene Perineura rubi og Tenthredopsis auriculata påtreffes her (figur 9). Et generelt trekk hos bladveps, er at de synes å foretrekke habitater med lite maur. Sannsynligvis er bladvepslarver spesielt utsatt for maurpredasjon.

-
- Lisens
- CC BY 3.0
- Opphav
- Ole Jørgen Lønnve
- Utgiver
- Biofokus
- Last ned (454.8 KB)
- Rediger (Krever innlogging)
Figur 7
Veikanter kan ha stor diversitet av planteveps. Langs slike kanter kan mange arter Tenthredo forekomme, men også flere arter Macrophya, Tenthredopsis, Rhogogaster og Aglaostigma. I tillegg forekommer en rekke andre planteveps i tilknytning til vegetasjonen langs veien. Fra Løten i Hedmark.

-
- Lisens
- CC BY 3.0
- Opphav
- Ole Jørgen Lønnve
- Utgiver
- Biofokus
- Last ned (486.1 KB)
- Rediger (Krever innlogging)
Figur 8
I bjørk- og vierregionen i fjellet forekommer flere arter Tenthredininae. Rhogogaster viridis og R. punctulata er vanlige her, mens Pachyprotasis rapae og flere arter Tenthredo også kan forekomme i slike habitater. I tillegg er underfamilien Nematinae særlig artsrik. Fra Øvre Heimdalsvann, Øystre Slidre i Oppland.

-
- Lisens
- CC BY 3.0
- Opphav
- Ole Jørgen Lønnve
- Utgiver
- Biofokus
- Last ned (6.5 MB)
- Rediger (Krever innlogging)
Figur 9
Mosaikkpregete habitater i tilknytning til vassdrag kan huse flere arter. Arter som Tenthredopsis auriculata, Perineura rubi, Macrophya albicincta samt flere Tenthredo-arter er tilknyttet slike habitater. Fra Ringnesbekken i Askim, Østfold.
Innsamling av planteveps
Planteveps samles best inn ved å gå og håve i vegetasjonen. Malaisefeller fungerer også bra (figur 10), men disse samler gjerne mer selektivt. Enkelte arter i underfamilien Tenthredininae går ikke veldig hyppig i malaisefeller, spesielt gjelder dette kanskje enkelte Tenthredo-arter. Selv om disse også forekommer i malaisefellemateriale, vil man som regel raskt få en mye bedre oversikt over hvilke arter som forekommer på en lokalitet ved å håve i vegetasjonen. For de artene som søker til blomster, er oppsøk av blomster en fin måte å få tak i arter på. Ofte kan man bare plukke dyrene med fingrene når de sitter på blomstene.
Planteveps bør nåles på insektnåler. Vingene og beina bør stå litt ut fra kroppen. På hunnene bør sagen vippes ut, men for underfamilien Tenthredininae lar bestemmelse, i hvert fall for nordisk materiale, som regel seg gjøre uten studering av sagen, og det er derfor ikke nødvendig å vippe den ut (som kan være litt fiklete å få til). Det samme gjelder de hannlige genitaliene. Som regel er det heller ikke nødvendig å gjøre dette, men hvis man gjør det, kan de limes opp på en liten plate og settes på nålen under dyret. Generelt bør voksne planteveps helst samles tørt, men mange arter, spesielt innen underfamilien Tenthredininae, tåler også godt å legges på sprit. Man må derimot være oppmerksom på at fargene kan falme eller forandre seg litt på dyr som har ligget lenge på sprit.
Ved innsamling av larver, er det viktig å notere vertsplante. Informasjon om vertsplanten til mange arter er mangelfull, slik at informasjon om dette er svært verdifull. Larver er dessverre vanskeligere å identifisere enn voksne dyr. En god ide er da å forsøke å klekke larvene, men dette kan være forholdsvis vanskelig, og man bør regne med å måtte eksperimentere litt. For bestemmelse av larver henvises til Lorentz & Kraus (1957).

-
- Lisens
- CC BY 3.0
- Opphav
- Ole Jørgen Lønnve
- Utgiver
- Biofokus
- Last ned (616.6 KB)
- Rediger (Krever innlogging)
Figur 10
Malaisefeller fanger forholdsvis bra med planteveps, men ikke alle arter går like lett i slike feller. Hvis man ønsker å kartlegge planteveps i et område bør malaisefeller derfor suppleres med andre metoder, spesielt manuell innsamling med håv.
Referanser
Blank SM og Ritzau C (1998). Die Tenthredopsini Deutschlands (Hymenoptera: Tenthredinidae). Pp. 227–246 in Taeger, A. & Blank. S. M. (eds): Pflanzenwespen Deutschlands (Hymenoptera, Symphyta). Kommentierte Bestandsaufnahme. 364 pp. Verlag Goecke & Evers, Keltern.
Blank S og Taeger A (2006). Taxonomy and Evolution of Tenthredo (Elinora) Species similar to T. dahlia and T. koehleri (Hymenoptera: Tenthredinidae). In Blank, S. M., Schmidt, S. & Taeger, A. (eds). 2006. Recent Sawfly Research: Synthesis and Prospects. 199−227.
Endrestøl A, Gammelmo Ø, Hansen LO, Lønnve OJ, Olberg S, Olsen KM og Aarvik L (2007). Registrering og overvåking av utvalgte insektarter i Oslo kommune III 2007. 33 pp. NHM-rapport.
Gould I og Bolton B (1988). The Hymenoptera. Oxford University Press. 332 pp.
Goulet, H. 1996. Revision of the Nearctic species of the arcuate group of the genus Tenthredo with notes on the higher classification of the Tenthredinini (Hymenoptera, Symphyta, Tenthredinidae). Contributions of the American Entomological Institute. 92 (2), 1−135.
Kiær H (1893). Indberetning om en i det sydlige Norge foretagen entomologisk Reise sommeren 1891. Enomologisk Tidskrift, Stockholm 1893, 225–234.
Liston AD (1980). Why sawflies of the Tenthredo arcuate-schaefferi complex (Hymenoptera, Tenthredinidae) visit flowers. Ann. Ent. Fenn. 46, 85−88.
Lorentz H og Kraus M (1957). Die Larvalsystematik der Blattwespen (Tenthredinoidea und Megalodontoidea). 1−339 pp. Berlin.
Taeger, A., Altenhofer, E., Blank, S. M., Jansen, E., Kraus, M., Pschorn-Walcher, H. & Ritzau, C. 1998. Kommentare zur Biologie, Verbreitung und Gefährdung der Pflanzenwespen Deutschlands (Hymenoptera, Symphyta). Pp. 49 – 136 in Taeger, A. & Blank. S. M. (Eds): Pflanzenwespen Deutschlands (Hymenoptera, Symphyta). Kommentierte Bestandsaufnahme. Verlag Goecke & Evers, Keltern.
Taeger A, Blank S og Liston AD (2010). World Catalog of Symphyta (Hymenoptera). Zootaxa 2580, 1–1064.
Totland Ø, Hovstad KA, Ødegaard F og Åström J (2013). Kunnskapsstatus for insektpollinering i Norge – betydningen av det komplekse samspillet mellom planter og insekter. Artsdatabanken, Norge. 74 s.
Viitasaari M (Ed.) (2002). Sawflies (Hymenoptera, Symphyta). Tremex Press Ttd. Helsinki. 1. 516 pp.

-
Aglaostigma aucupariae
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (552.1 KB)
- Rediger (Krever innlogging)
Aglaostigma aucupariae hunn. Fyledalen, Sjöbo, Skånes län, Sverige. 22.05.2012. Ole J. Lønnve.

-
Macrophya montana
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (508.9 KB)
- Rediger (Krever innlogging)
Macrophya montana hunn. Nordåsen, Birkenes, Aust-Agder, Norge. Juni 2008. Leg. Svein Svendsen.

-
Pachyprotasis antennata
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (524.6 KB)
- Rediger (Krever innlogging)
Pachyprotasis antennata hunn. Grondo i Råsdalen, Lærdal, Sogn og Fjordane, Norge. 04.07.2013. Leg. Kai Berggren.

-
Perineura rubi
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (502 KB)
- Rediger (Krever innlogging)
Perineura rubi hunn. Åranstorpet, Kongsvinger, Hedmark, Norge. 16.05-14.06.2004. Leg Karsten Sund.

-
Rhogogaster punctulata
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (472.8 KB)
- Rediger (Krever innlogging)
Rhogogaster punctulata hunn. Bøkkestølen, Vang, Oppland, Norge. 30.06.2011. Leg. Kai Berggren.

-
Tenthredo mesomela
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (519.9 KB)
- Rediger (Krever innlogging)
Tenthredo mesomelas hunn. Midtre Ljøsne, Lærdal, Sogn og Fjordane, Norge. 30.06.2013. Leg. Kai Berggren.

-
Tenthredopsis scutellaris
-
- Lisens
- CC BY 3.0
- Opphav
- Kim Abel
- Utgiver
- Biofokus
- Last ned (416.6 KB)
- Rediger (Krever innlogging)
Tenthredopsis scutellaris hunn. Vennisviki, Vang, Oppland, Norge. 28.06.2013. Leg. Kai Berggren.